


 扫码安装网站APP(Android版)
扫码安装网站APP(Android版)
来源公众号:地球玩家2号
深度解析 · 细胞的衰老
细胞为什么会衰老?——从”端粒变短”到”末端失去保护
染色体末端如何避免被当作 DNA 断裂:T‑loop 与 shelterin
教材里的”端粒学说”只讲了一件事:端粒会随细胞分裂而缩短。但有一个被它略过的问题:染色体的末端,本身就是一个”游离的 DNA 末端”。而细胞内的损伤监测系统,恰恰把”游离末端”判定为最危险的损伤之一。那么,为什么正常染色体的末端没有天天触发警报、没有被错误地连接起来?本文从这个问题出发,把”端粒缩短”重新理解为一套末端保护结构的瓦解。
PART 01
末端识别问题:染色体的末端,为什么是个麻烦
DNA 双链断裂(double‑strand break,DSB)是细胞中最危险的损伤类型之一。一旦出现游离的双链末端,细胞会立即启动两类应对:先由损伤监测系统识别并发出信号,再由修复系统处理——例如把两个断裂端直接连接起来(非同源末端连接,NHEJ)。这套机制对修复真正的断裂是必要的。
问题在于:一条线性染色体的正常末端,在结构上和一个 DSB 几乎一样——都是一段没有”后续序列”的游离 DNA 末端。如果损伤监测系统”看到”了它,就可能把它当成断裂来”修复”:把两条不同染色体的末端连接到一起,造成染色体融合。融合的染色体在下一次分裂时会被纺锤体拉向两极并再次断裂,引发更严重的基因组混乱。端粒的首要任务,正是让末端不被识别为断裂,学术上称为末端保护问题(end‑protection problem)。
PART 02
端粒的真实结构:不止是一段重复序列
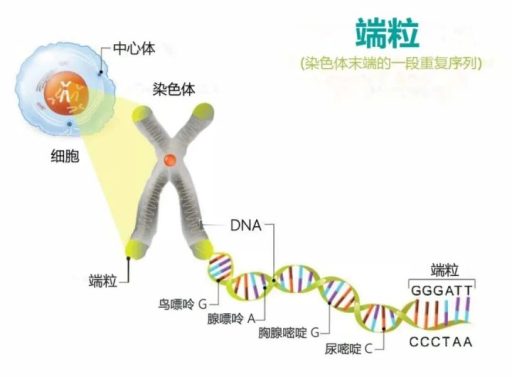
高中阶段把端粒理解为”染色体末端的一段特殊 DNA 序列”,这没有错,但不完整。人类端粒的实际构成是:
- 双链重复区:以 5′‑TTAGGG‑3′ 为单位的串联重复,人类约为 2–15 kb(千碱基对)。
- 3′ 单链突出:末端有一段富含 G 的单链伸出(约 50–300 个核苷酸),即”G‑突出”。它是后面整个保护结构的关键。
- 结合蛋白:一组专门识别端粒序列的蛋白结合其上,与 DNA 共同构成动态的”末端保护复合体”。
换句话说,端粒不是一段裸露的 DNA,而是“特定 DNA 序列 + 专属蛋白”组成的一套结构。它如何让一个游离末端”不像游离末端”?答案分两层:先看 DNA 自身的折叠方式,再看蛋白的作用。
PART 03
T‑loop:把末端隔离在双链内部
1999 年,Griffith 与 de Lange 两个实验室用电子显微镜观察到一个出人意料的结果:哺乳动物端粒的末端并不是一条伸直的线,而是回折成一个大的环状结构,称为 T‑loop(端粒环)。
其几何机制是:末端的 3′ 单链突出回折,插入到上游的双链端粒重复区中,与其中的互补链(C 链)配对,把原来的同序列链(G 链)局部顶开,形成一个小的置换环(D‑loop)。这样一来原本暴露在外的游离末端,被”塞回”双链内部,不再以一个可被识别的自由 DNA 末端的形式存在。
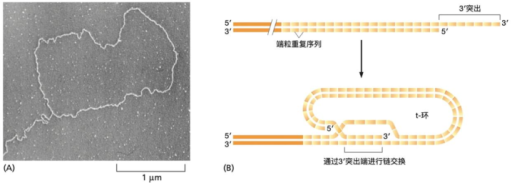
这是一个纯粹的结构性解决方案:末端没有被改造,也没有被”修复”,只是被折叠隐藏,使损伤监测系统”无从识别”。但 DNA 自己不会无缘无故折成这样,T‑loop 的形成与维持,依赖端粒上的一组蛋白。
PART 04
shelterin:双链与单链的两道防线(拓展,可不看)
2005 年,de Lange 把结合在端粒上的六个蛋白统称为 shelterin(庇护蛋白复合体),成员是 TRF1、TRF2、RAP1、TIN2、TPP1、POT1。它们各司其职,但对理解”末端保护”最关键的是两个直接的执行者,分别守住双链区和单链突出。双链端粒重复(TTAGGG)ₙ3′ 单链突出TRF1TRF2POT1TIN2TPP1RAP1针对”双链末端”的探测(ATM 信号 / 末端连接 NHEJ)被 T‑loop + TRF2 抑制针对”单链 DNA”的探测(ATR 信号)被 POT1 遮盖突出而抑制两套损伤探测系统 ↔ 两道端粒防线图 3 shelterin 的分工。TRF1、TRF2 结合双链区,POT1 结合单链突出,TIN2/TPP1/RAP1 起桥接作用。细胞有两套末端探测系统,端粒以两道防线分别应对。
TRF2 —— 隐藏双链末端。它结合双链端粒重复,是 T‑loop 形成与维持所必需的。它的作用是抑制针对”双链末端”的损伤信号(ATM 通路)与末端连接修复(NHEJ)。实验中过表达失活型 TRF2,会导致端粒失去保护、出现染色体末端融合——这从反面证明了它对末端的保护作用。
POT1 —— 遮盖单链突出。它特异结合 3′ 单链突出,遮盖这段单链 DNA,阻止单链探测因子(RPA)结合,从而抑制针对”单链 DNA”的损伤信号(ATR 通路)。
为何要两道防线? 因为细胞本就有两套独立的损伤探测:一套盯”双链断裂末端”,一套盯”暴露的单链 DNA”。端粒末端两者皆有,于是 TRF2(连同 T‑loop)负责让双链末端不被识别,POT1 负责遮盖单链突出。其余成员中,TIN2、TPP1 负责把 POT1 连接到复合体上,TRF1 主要参与端粒区的正常复制,而非直接参与末端保护。
PART 05
重新理解”端粒学说”:长度为结构服务
由于末端复制问题(线性 DNA 复制时末端无法被完整复制),体细胞每分裂一次,端粒就缩短一段(约几十至一两百个碱基对)。教材到这里通常就停了,结论是”端粒变短 → 细胞衰老”。但“变短”为什么会导致衰老,真正的关键在结构:长端粒T‑loop 稳定多次分裂逐渐缩短临界长度约 2–4 kbT‑loop 无法维持!末端暴露触发损伤应答细胞衰老(停止分裂)/ 凋亡”长度变短”的真正后果,是末端保护结构失效衰老不是因为”DNA 用完了”,而是因为末端不再能被有效保护、开始被当作损伤图 4 从”长度”到”结构”再到”衰老”的因果链。端粒短到临界长度时,没有足够的重复序列来形成稳定的 T‑loop、容纳 shelterin,末端保护随之瓦解。
当端粒短到某个临界长度(约 2–4 kb)时,已没有足够的重复序列来折叠出稳定的 T‑loop、也难以容纳完整的 shelterin。末端保护一旦失效,末端就被当作损伤,触发 DNA 损伤应答,细胞随之进入衰老(不可逆地停止分裂)或走向凋亡。
所以本文想强调的是:端粒缩短之所以重要,不是因为”长度”本身,而是因为长度是结构保护的物质基础。端粒缩短,本质上是一套末端保护系统的逐步瓦解。
PART 06
端粒酶与癌细胞:
既然端粒会缩短,为什么生殖细胞、干细胞乃至癌细胞能长期维持端粒?答案是端粒酶。
关键区分:端粒酶的职能是”补长度“——它是一种核糖核蛋白酶,含逆转录酶亚基(TERT)和一段 RNA 模板(TERC),以自身 RNA 为模板,把 TTAGGG 重复合成到染色体的 3′ 末端,补回缩短的部分。而”搭结构“(折叠 T‑loop、容纳 shelterin、屏蔽末端)是端粒 DNA 与 shelterin 蛋白完成的。两者配合,但分工不同:端粒酶不直接形成保护结构,它提供的是结构得以搭建的”原料长度“。端粒酶如何”补长度”新合成的 TTAGGG端粒酶TERT + RNA 模板同一支酶,两种结局正常体细胞端粒酶活性低 / 几乎无→ 端粒随分裂持续缩短→ 保护失效 → 衰老作为一道天然的抗癌屏障癌细胞重新激活端粒酶(少数走 ALT)→ 维持端粒长度与末端保护→ 越过衰老屏障获得近乎无限的增殖能力图 5 端粒酶以自身 RNA 为模板合成端粒重复序列。约 85–90% 的癌细胞通过重新激活端粒酶来维持端粒,这正是教材”癌细胞能够无限增殖”的一个分子层面解释。
在大多数正常体细胞中,端粒酶活性很低甚至缺失,端粒随分裂缩短,最终触发衰老。这其实是一道天然的抗癌屏障:限制了细胞的分裂次数。而约 85–90% 的癌细胞会重新激活端粒酶(少数通过另一条名为 ALT 的途径),维持端粒长度,从而维持末端保护、越过衰老限制,获得近乎无限的增殖能力。
小结:一次认知升级
把这条线索串起来,对端粒的理解就从”一段会变短的 DNA”升级为一套动态的末端保护系统:
- 线性染色体的末端先天像一个 DNA 双链断裂,若不处理就会被错误”修复”(融合);
- 端粒用 T‑loop 把末端折叠隐藏,用 shelterin(TRF2 作用于双链、POT1 遮盖单链)抑制两套损伤探测;
- 端粒缩短的危害,本质是保护结构的瓦解——长度是为结构服务的;
- 端粒酶负责”补长度”,与”搭结构”的 shelterin 系统分工不同;癌细胞重新激活端粒酶以维持这套保护。
📐 知识分层(请按需取用)
高中核心(考查范围):端粒位于染色体末端、随细胞分裂而缩短;端粒学说与细胞衰老的关系;端粒酶能延长端粒,使生殖细胞、癌细胞等维持增殖能力。
拓展延伸(不作考查要求):末端保护问题、T‑loop 与置换环(D‑loop)、shelterin 复合体(TRF2/POT1 等)、ATM/ATR 两套损伤探测通路、ALT 途径。这些属于大学与科研层面内容。
注:文中端粒长度(约 2–15 kb 双链 + 50–300 nt 单链突出)、临界长度(约 2–4 kb)、缩短幅度(每次约数十至一两百 bp)及癌细胞端粒酶激活比例(约 85–90%)为不同文献的常见取值范围,个体与细胞类型间存在差异。图中比例经简化处理。



近期评论