

 扫码安装网站APP(Android版)
扫码安装网站APP(Android版)
来源公众号:生命教育观察 作者:小强大生物
教材在介绍体液免疫的基本过程时,相关信息里介绍这样的一段话:通常情况下,一个B细胞只针对一种特异的病原体,活化、增殖后只产生一种特异性的抗体。人体内B细胞的种数在 10⁹以上,至少可以产生10⁹种独特的抗体,可以识别自然界存在的种类繁多的病原体。
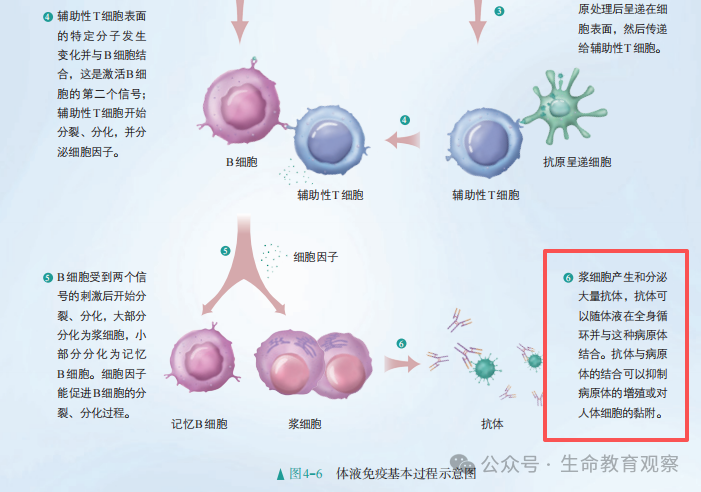
如何去理解抗体种类的多样性,这种多样性又是如何产生的呢?
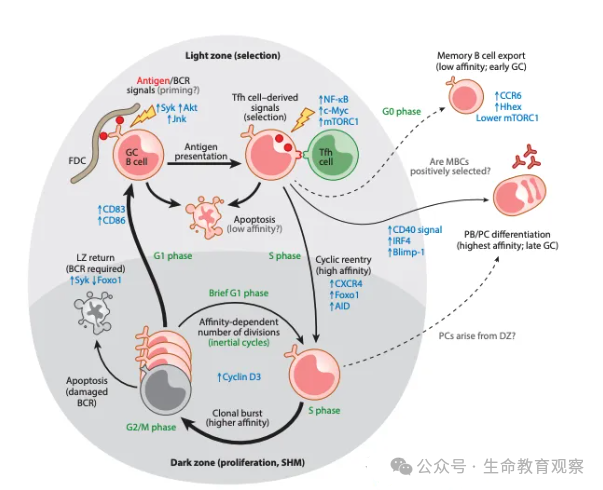
抗体的多样性核心源于V (D) J 重组机制,免疫系统通过基因片段的随机组合、精准修饰,让有限的基因片段产生海量特异性抗体。
抗体基因是拆分的片段库
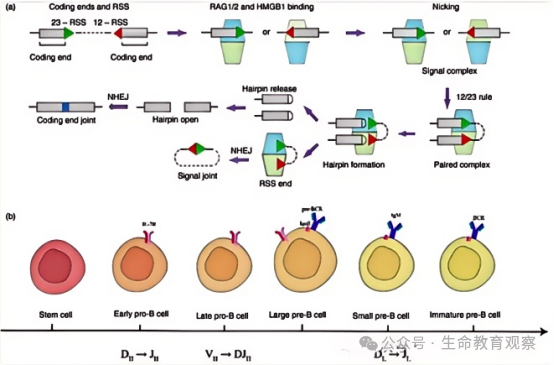
人体基因组中,负责编码抗体(B 细胞受体,BCR)的基因并非完整序列,而是被拆分为三类可重组的片段。V 片段(可变区):决定抗体识别抗原的核心区域,数量成百上千;D 片段(多样性区):仅存在于抗体重链基因中,进一步增加序列多样性;J 片段(连接区):连接 V/D 片段与恒定区,参与抗体结构组装。每一个新生 B 细胞发育时,会从这些片段库中 “随机抽取” 特定 V、D、J 片段,通过 DNA 剪切与拼接组装成完整的抗体可变区基因 —— 这是多样性产生的 “初始框架”。
两类关键分子的协作分工重组执行
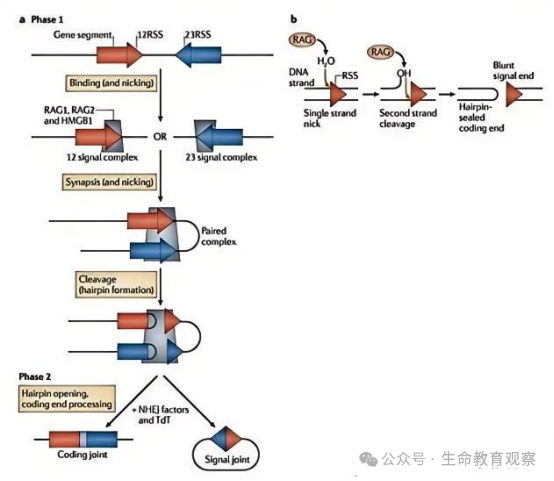
V (D) J 重组的实现依赖两类核心分子,如同 “工人 + 修理队” 的配合。
切割工人:RAG1/RAG2 蛋白复合物需在HMGB1 蛋白辅助下发挥作用,核心功能是识别并切割 DNA。先识别片段两侧的重组信号序列(RSS)(由 “9bp 非保守序列 + 12/23bp 间隔区 + 7bp 保守序列” 组成,分为 12RSS 和 23RSS);遵循12-23 规则:仅催化 “1 个 12RSS+1 个 23RSS” 配对,避免片段错误连接;切割过程:先在 RSS 邻近 DNA 形成单链切口,释放 3′-OH 末端;该末端进一步攻击对链磷酸二酯键,形成双链断裂,最终产生 “发夹结构的编码末端”(V/D/J 片段侧)和 “平端的信号末端”(RSS 侧)。
连接修理队:非同源末端连接(NHEJ)机制负责将断裂的 DNA 重新连接,且对 “编码末端” 进行关键修饰 —— 这是多样性升级的核心步骤。编码末端会发生随机核苷酸缺失,改变片段连接处的序列;同时,末端脱氧核苷酸转移酶(TdT) 会随机添加 “非模板核苷酸(N 区)”,进一步延长并丰富连接区序列;最终形成 “编码连接”(修饰后的 V-D-J 片段连接),而信号末端则未经修饰直接形成 “信号连接”,确保基因结构稳定。
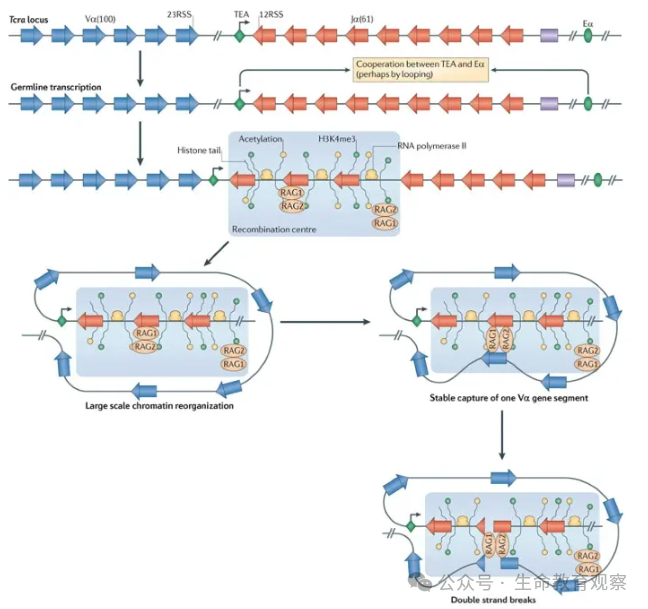
染色质动态确保重组精准高效调控机制
V (D) J 重组并非随机发生,需依赖染色质层面的调控,确保片段可及性与重组特异性。
重组中心的形成抗体基因座(如重链、轻链基因区)会通过DNA 环化,让远端的 V 片段与近端的 D/J 片段物理靠近;同时,TEA 启动子与Eα 增强子协同激活 “种系转录”,在染色质上建立 “开放标记”(如组蛋白乙酰化、H3K4me3 修饰、RNA 聚合酶 II 富集),为 RAG 蛋白招募创造条件。
逐步解放模型应对 RSS 可及性问题部分 RSS(如 23RSS)常被核小体包裹,导致其 7bp 保守序列难以被 RAG 识别,需通过时序协调解决。第一步:RAG1/RAG2-HMGB1 先结合 23RSS 的 9bp 区域,再与 12RSS 配对;第二步:RAG 先切割 12RSS 的 7bp 区域,触发染色质动态重塑;第三步:23RSS 的 7bp 区域从核小体表面暴露,RAG 完成切割;最终:配对复合物形成发夹结构,确保双链断裂精准发生。
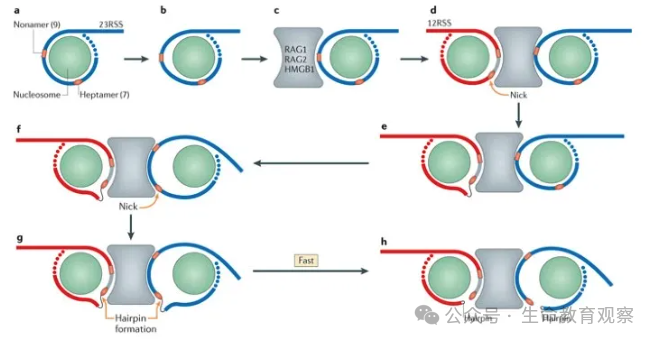
多样性的叠加效应
上述过程通过三重机制叠加,产生海量抗体。片段随机组合成百上千的 V、D、J 片段随机抽取,仅组合方式就可产生数万种序列;编码末端修饰核苷酸缺失 + TdT 添加 N 区,让每一次连接都产生独特序列;染色质调控下的精准重组,确保片段连接的特异性,避免无效重组,同时扩大可重组片段的范围。最终,这套机制让免疫系统能产生10¹¹ 数量级的特异性抗体,足以识别几乎所有可能的外来抗原(如病毒、细菌),实现对病原体的精准防御。
来源网址:精读教材:抗体的多样性是怎么产生的?


近期评论