

 扫码安装网站APP(Android版)
扫码安装网站APP(Android版)
来源公众号:久劳斯的生物圈 作者:汪久佳
【说明】多年前的一次面试中,我抽到的课题是“DNA的复制”。说完课后,评委问我:“你对梅塞尔森和斯塔尔这两位科学家了解多少?”我一时不知如何回答,只能如实说,除了教材上的内容,我对他们并无更多了解。当时不免有些尴尬,背后直冒冷汗。最近查阅文献,才大体了解了这段科学史。在此将文字稍作整理,聊作记录。如有谬误,敬请指正。
1958年,马修·梅塞尔森(Matthew Meselson)与富兰克林·斯塔尔(Franklin Stahl)在《美国国家科学院院刊》(PNAS)上发表了一篇短文,题为《DNA复制的机制》。这篇看似简洁的文章,却一举解决了分子生物学诞生初期最核心、最富争议的难题之一——DNA如何复制自身。他们的实验,被后世誉为“生物学中最美丽的实验”或“最优雅的实验”。本报告旨在超越对实验步骤的简单复述,深入追溯其发现的历史脉络,剖析其背后的科学思想革命,详细解构其精妙绝伦的实验设计,并全面评估其对现代分子生物学乃至整个生命科学所产生的深远影响。我们将看到,Meselson-Stahl实验不仅是一个技术上的杰作,更是一场逻辑、美学与科学哲学完美结合的典范,它在一个关键的历史节点,为遗传信息传递的物理机制提供了决定性的、直观的证明,从而稳固了分子生物学大厦的基石。
1 黎明前的迷雾——DNA复制问题的提出与理论争鸣(1953–1957)
要理解Meselson-Stahl实验的伟大与必要性,我们必须首先回到20世纪50年代初那个激动人心又充满不确定性的科学图景中。
1.1 双螺旋的震撼与一个未经验证的推论
1953年4月,詹姆斯·沃森(James Watson)和弗朗西斯·克里克(Francis Crick)在《自然》杂志上发表了那篇划时代的短文,提出了DNA的双螺旋结构模型。这个模型的美学与逻辑力量立即征服了科学界。它不仅解释了X射线衍射数据,更重要的是,它以一种极其直观的方式暗示了遗传物质可能的功能机制:碱基对序列承载遗传信息,而两条链的互补性则暗示了复制潜力。
在论文接近结尾处,沃森和克里克写下了一句充满洞察力却又小心翼翼的话:“我们并未忽视,我们所假设的特定配对方式,立即暗示了遗传物质一种可能的复制机制。”他们随后简要描述了一种机制:双螺旋解开,每条链作为模板,按照碱基互补规则合成一条新的互补链。最终,一个DNA分子产生两个完全相同的子代分子,每个都包含一条来自亲代的“旧”链和一条新合成的链。他们将这种模式称为“半保留复制”。
然而,必须强调,这仅仅是一个基于结构模型的推论,而非实验证明。在当时,这甚至是整个模型中最脆弱、最需要验证的环节之一。双螺旋结构本身有罗莎琳德·富兰克林和莫里斯·威尔金斯的X射线衍射数据强力支持,但复制机制纯粹是理论猜想。沃森和克里克本人也深知这一点,他们的表述充满了试探性。
1.2 三种假说的对峙
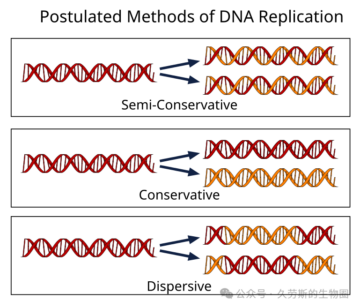
很快,科学界围绕DNA复制可能的具体模式展开了激烈的讨论和猜想。综合起来,主要形成了三种相互竞争的理论模型:
1.半保留复制:即沃森-克里克模型。复制后,每个子代双链DNA分子由一条亲代链和一条新合成的子链组成。这是最符合双螺旋互补结构直觉的模型。
2.全保留复制:亲代DNA双链在复制过程中保持完整,作为一个整体模板,指导合成一个全新的、完全由新链组成的子代DNA双链。复制后,一个分子是“全旧”的,另一个是“全新”的。这种模型在一些科学家看来似乎更“经济”或更“简单”,因为它避免了复杂的链分离问题。
3.分散复制:这是三种模型中最复杂、也最难以直观想象的一种。它认为亲代DNA双链在复制时会被打成片段,新旧片段随机地混合在一起,共同构成两个子代分子。因此,每个子代分子的每一条链都是新旧DNA片段拼接而成的“马赛克”。这种模型由马克斯·德尔布吕克等人提出,部分是为了解释当时认为的DNA复制可能面临的拓扑学难题。
这三种模型在理论上都能解释DNA的倍增,但预言了完全不同的子代DNA分子物理组成。在1950年代中期,没有任何技术能够直接观测或区分这些差异。因此,DNA复制机制成了一个悬而未决的核心争议,直接关系到双螺旋模型的核心预言是否正确,以及遗传信息传递的基本原理究竟是什么。
1.3 早期探索与困境
在Meselson和Stahl之前,已有科学家尝试解决这个问题。其中最著名的是1957年由J. Herbert Taylor、Philip Woods和Walter Hughes利用植物根尖细胞和放射性同位素标记进行的实验。他们通过放射自显影观察染色体复制,发现复制后的染色体中,两条染色单体一条标记信号强,一条弱,这被解读为支持半保留复制。然而,这个实验系统存在局限性:它观察的是宏观的染色体水平,而非单个DNA分子;放射性标记和自显影技术分辨率有限,解释上存在模糊性;更重要的是,它无法严格排除分散复制的可能性。因此,泰勒的实验虽然极具启发性,但未能一锤定音,争议依然存在。
科学界急需一个更直接、更清晰、能在分子水平上区分三种模型的实验。这个实验需要满足几个苛刻条件:第一,必须能追踪亲代DNA的物理去向;第二,必须能高分辨率地区分不同组成的DNA分子;第三,实验设计必须逻辑严密,能给出无歧义的预测结果。正是在这样的背景下,两位年轻的科学家——梅塞尔森和斯塔尔,在加州理工学院的学术沃土上,开始了他们的合作。
2. 思想的熔炉——Meselson与Stahl的相遇与实验思想的诞生
伟大的实验往往源于伟大的思想碰撞,以及孕育这些思想的独特环境。Meselson-Stahl实验的诞生,深深植根于20世纪50年代加州理工学院分子生物学摇篮的独特氛围中。
2.1 两位主角:知识背景与结合
马修·梅塞尔森:当时是加州理工学院的研究生,师从著名的化学家莱纳斯·鲍林。梅塞尔森拥有深厚的物理化学背景,精通于同位素和超速离心技术。他思维严谨,对实验的精确性和逻辑的优雅性有着近乎偏执的追求。他最初感兴趣的问题是蛋白质合成,但在接触到DNA复制问题后,被其逻辑清晰性所吸引。
富兰克林·斯塔尔:作为一名博士后,斯塔尔拥有遗传学背景,曾与噬菌体遗传学先驱之一萨尔瓦多·卢里亚共事。他擅长遗传学思维和生物学问题的构思,对DNA复制这个核心的遗传学问题有着天然的兴趣。
两人的结合堪称完美:梅塞尔森带来了解决难题所需的尖端物理化学工具和严谨的实验设计思维,而斯塔尔则确保了整个研究牢牢锚定在最具生物学意义的核心问题上。这种跨学科的互补性,是实验成功的关键因素之一。
2.2 学术环境:加州理工学院的“噬菌体小组”氛围
加州理工学院在当时是分子生物学革命的中心之一。马克斯·德尔布吕克在这里建立了著名的“噬菌体小组”,吸引了沃森、克里克等一大批顶尖头脑。这里的学术氛围极其自由、开放且充满活力,鼓励大胆的猜想、激烈的辩论和跨领域的合作。年轻的科学家被鼓励去挑战最根本的问题,而不必拘泥于传统的学科界限。
这种环境对梅塞尔森和斯塔尔产生了深远影响。他们可以随时与德尔布吕克(分散复制模型的提出者)、沃森、克里克等人交流想法,接受最直接的批评和挑战。正是与德尔布吕克的讨论,促使他们必须设计一个能够同时、明确地区分所有三种模型的实验,而不仅仅是证明半保留复制。这种高标准的科学要求,最终催生了实验无与伦比的逻辑严密性。
2.3 核心灵感的迸发:从“重氮”到“密度梯度离心”
实验的核心技术灵感来源于多个领域的融合。
首先,同位素标记的选择。梅塞尔森和斯塔尔需要一种方式来“标记”亲代DNA,使其在复制后能被追踪。他们选择了氮元素,因为它是DNA碱基的核心组成元素。更妙的是,氮有两种稳定的同位素:常见的轻氮(¹⁴N)和较重的重氮(¹⁵N)。¹⁵N没有放射性,但原子质量比¹⁴N约重6.7%。如果让生物在富含¹⁵N的培养基中生长多代,其DNA中的所有氮原子都将被¹⁵N取代,形成“重”DNA。将其转移到普通的¹⁴N培养基中后,新合成的DNA链将使用¹⁴N,形成“轻”链。关键在于,¹⁵N和¹⁴N的化学性质几乎完全相同,不会干扰正常的生物学过程,唯一区别就是质量。这是一个完美的“寂静标记”。
其次,也是最关键的突破,是如何分离和检测这些质量有细微差别的DNA分子。传统的沉降速度离心无法区分。这时,梅塞尔森想到了当时一项较新的技术——平衡密度梯度离心,特别是使用氯化铯溶液的方法。这项技术由生物物理学家杰罗姆·维诺格拉德等在加州理工学院开发。其原理是:在高转速下,CsCl盐离子在离心力场中形成从管底到管顶的连续密度梯度。当DNA分子被加入其中并长时间离心时,它们会迁移到与其自身浮力密度相等的CsCl密度区域,并聚集形成一条狭窄的条带。条带的位置可以通过紫外吸收照相来精确定位。
梅塞尔森的天才之处在于,他意识到¹⁵N标记的“重”DNA和¹⁴N组成的“轻”DNA,其浮力密度存在可测量的差异。更重要的是,如果复制是半保留的,第一代子代DNA将是一条重链和一条轻链组成的杂交分子,其密度应该恰好介于重DNA和轻DNA之间。而全保留复制预言第一代产物是一个全重分子和一个全轻分子,应出现两条分离的带。分散复制则预言所有子代DNA的密度都相同,且介于轻重之间,但随着复制代数增加,这条“中带”会逐渐变轻,而不会产生离散的轻带。
至此,一个清晰、可检验、可区分的实验蓝图在脑海中形成。这个设计的美在于,它不仅仅是一个“尝试看看”的实验,而是一个基于严密演绎推理的“判决性实验”。他们首先从三种理论模型出发,推导出各自对DNA密度分布在不同复制代数后的明确预测,然后设计实验去检验这些预测。这是一种假设-演绎法的经典应用,充满了科学的理性之美。
3. 优雅的判决——实验设计、执行与数据的直接解读
让我们深入细节,重现这个“最美丽实验”的具体实施过程,并欣赏其数据呈现的清晰与力量。
3.1 实验材料与培养策略
1.模式生物选择:他们选择了大肠杆菌 B菌株。这是一个标准且易于操作的微生物模型,生长快速,遗传背景清晰。
2.同位素标记:制备以¹⁵N为唯一氮源的“重”培养基,以及普通的以¹⁴N为氮源的“轻”培养基。
3.培养流程:
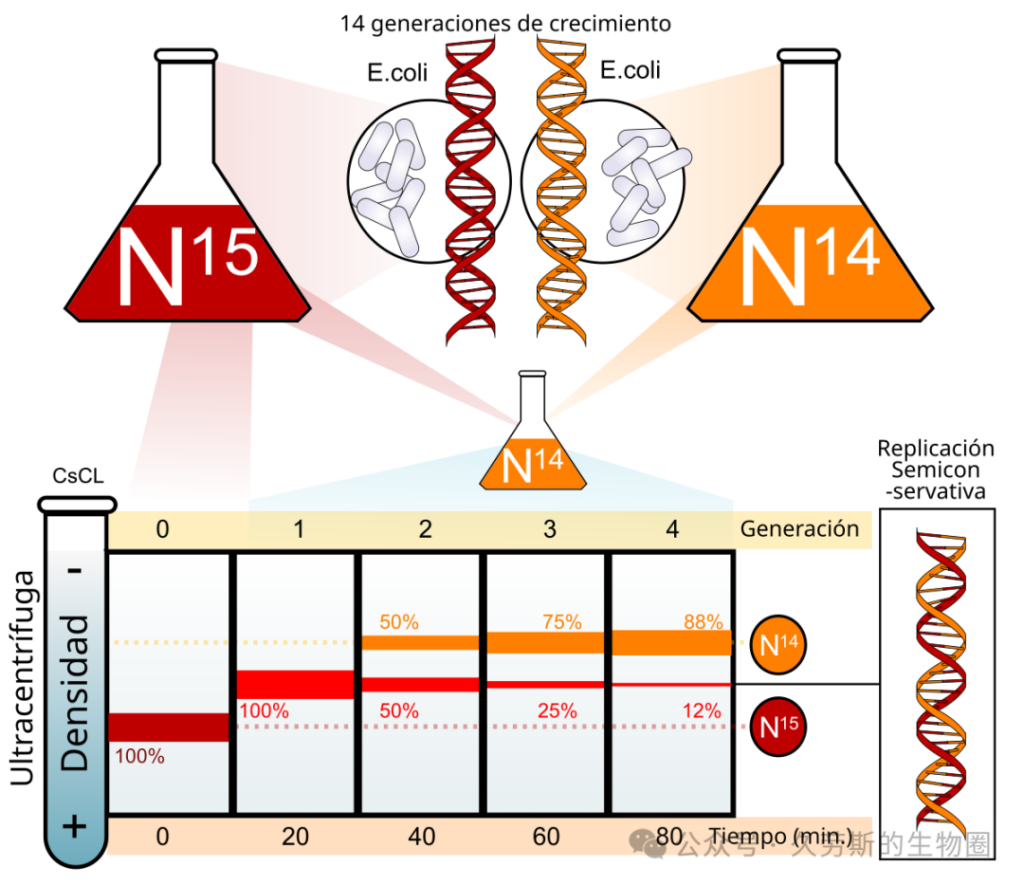
- 步骤一:制备“重”DNA。将大肠杆菌在¹⁵N培养基中连续培养约14代。理论上,经过这么多代生长,几乎所有氮原子都被¹⁵N替换,DNA达到均匀的“重”状态。
- 步骤二:世代零取样。取一部分细菌,提取其DNA,进行CsCl密度梯度离心分析。预期看到一条高密度的“重”带。
- 步骤三:转移与同步复制。将剩余的“重”细菌快速洗涤并转移到新鲜的、富含营养的¹⁴N培养基中。此时,细菌开始以¹⁴N为原料合成新的DNA。通过控制营养和生长条件,他们使细菌群体开始基本同步的复制。
- 步骤四:按时间/世代取样。在转移后的不同时间点,取样细菌,提取DNA。
3.2 核心技术:CsCl密度梯度离心
这是整个实验的“眼睛”。他们将提取的DNA与浓CsCl溶液混合,注入离心管,在分析型超速离心机中以约44,770 rpm的速度离心长达20小时以上。在离心过程中,CsCl形成稳定的密度梯度。DNA分子在离心力和扩散力的平衡下,最终在与其浮力密度匹配的位置聚集形成狭窄的条带。
他们采用了一种巧妙的紫外吸收照相技术来可视化条带:离心机配备了一个光学系统,用紫外光扫描离心管,并将吸收信号记录在照相底片上。吸收越强的位置,DNA浓度越高,在底片上显示为越黑的条带。这种方法可以精确定量条带的位置和强度。
3.3 决定性的数据与一目了然的解读
1958年《美国国家科学院院刊》上那篇简洁的论文,随文所附的几张实验图像,以其无可辩驳的清晰性成为了科学史上的经典。这些图像并非简单的示意图,而是实验本身的直接记录,每一张都对应着判决的关键一步。
技术的验证:区分轻重DNA的能力
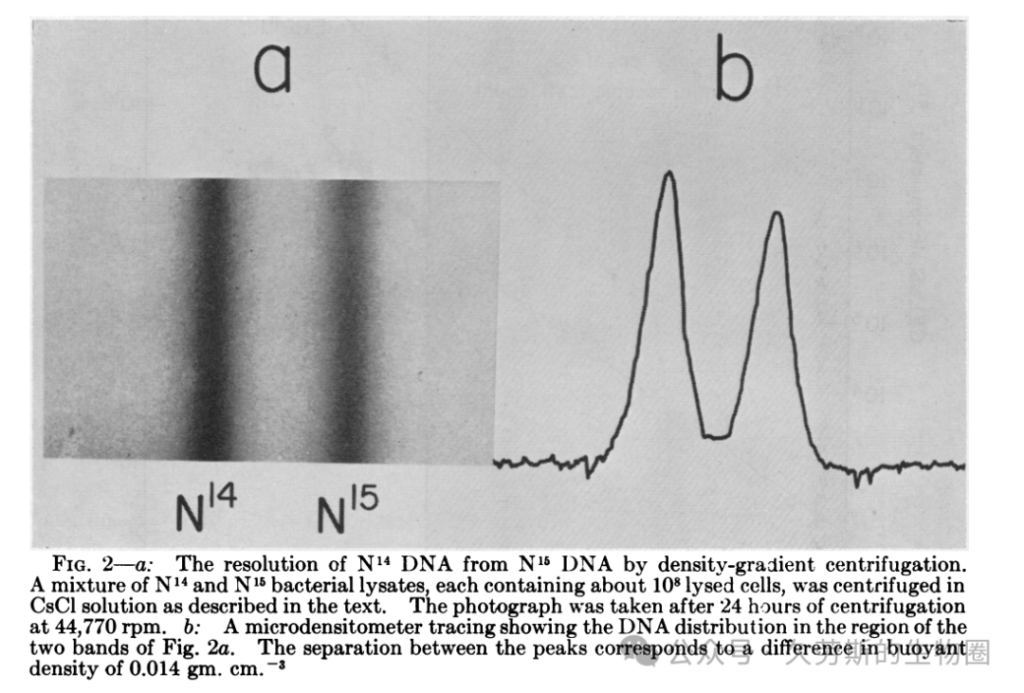
首先,实验必须证明其所依托的技术——氯化铯密度梯度离心——足以分辨由¹⁵N(重氮)和¹⁴N(轻氮)构成的DNA之间微小的密度差异。论文中的一张图示(通常被教科书引用为关键证据之一)展示了将分别培养的“重”DNA(¹⁵N)和“轻”DNA(¹⁴N)混合后离心的结果。紫外吸收照片上,两条清晰分离的条带出现在不同的密度位置,直观地证实了该方法拥有将两者区分开来的分辨率。这是整个实验逻辑得以成立的技术基石。
第一代:排除“全保留复制”
当将长期在¹⁵N培养基中培养的细菌转移到¹⁴N培养基中,并让其生长大约一个世代的时间后,对DNA进行分析,得到了一个决定性结果。离心图像显示,此时所有的DNA形成 单一的一条条带,而这条带的位置既不在原先重DNA的位置,也不在轻DNA的位置,而是精确地位于两者正中间。
这一现象具有 immediate 的判决意义:它直接排除了“全保留复制”模型。因为如果复制是全保留的,亲代完整的“重”DNA双链应当原封不动地保留,同时产生全新的“轻”DNA双链,那么在离心结果中理应同时观察到分离的“重带”和“轻带”。实验中仅出现的“中带”否定了这一图景。此时,实验结果与“半保留复制”和“分散复制”在第一代的预测是相符的,两者都预期产生密度居中的杂交分子。
第二代及以后:在“半保留”与“分散”之间做出裁决
真正的胜负在于后续世代。当细菌在¹⁴N培养基中生长约两个世代后,取样分析的结果呈现出第二个决定性图景:紫外吸收照片上出现了两条清晰可辨的条带。一条仍然位于第一代出现的中间密度位置,另一条则位于纯轻DNA(¹⁴N)的位置。随着世代继续增加,中间密度条带的强度保持相对稳定,而轻DNA条带的强度则持续增强。
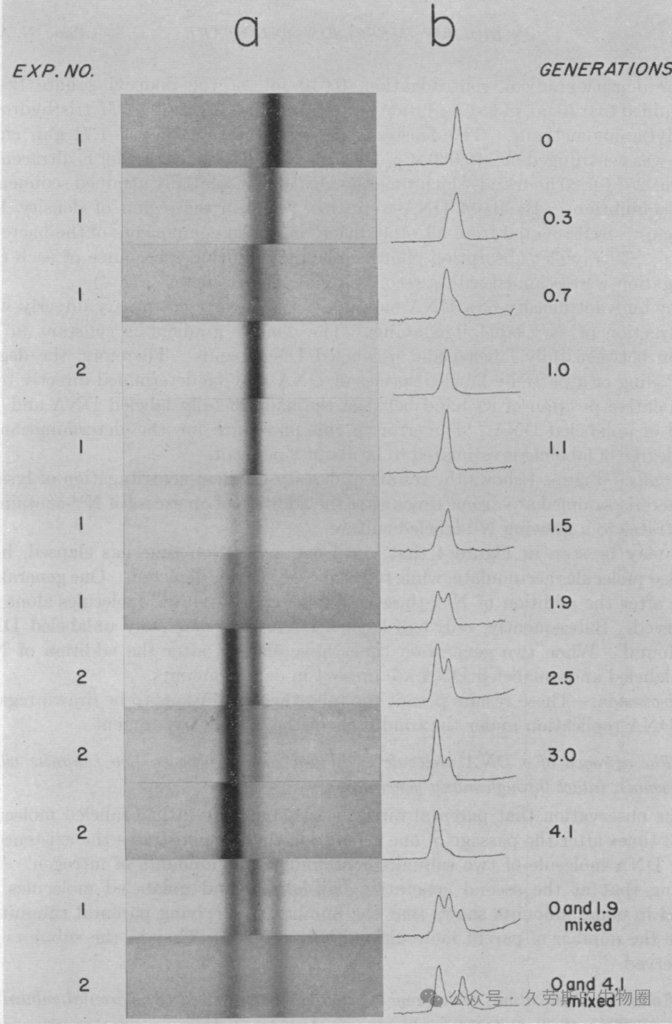
这一结果完美且独占地符合“半保留复制”模型的预测:第一代产生的杂交分子(一条¹⁵N链,一条¹⁴N链)在第二轮复制中,每条链分别作为模板。其中¹⁵N“重”链与新的¹⁴N链结合,再次形成密度居中的杂交分子;而¹⁴N“轻”链与另一条新的¹⁴N链结合,则形成密度全轻的分子。因此,第二代产物正是中密度与轻密度分子各占一半的混合物,表现为两条分离的条带。
与此同时,这一结果彻底证伪了“分散复制”模型。因为分散复制预言,亲代的¹⁵N原子会随着复制代数的增加,被不断稀释和分散到所有子代分子中。其预期结果不会是形成稳定、离散的中带和轻带,而是所有DNA分子的密度呈现为一个从中间密度向轻密度逐渐、连续移动的单一宽峰或弥散带。实验中出现的两条界限分明的条带,与分散复制的预言截然不同。
至此,Meselson和Stahl通过一组逻辑严密的序列图像,不仅正面验证了半保留复制,更以“排除法”这一强有力的科学逻辑,逐一否定了与之竞争的其他假说。图像本身,便是结论。
3.4 逻辑的终局:无可辩驳的证明
通过追踪从0代到4代的密度分布变化,Meselson和Stahl呈现了一幅动态的、符合严格逻辑推演的图景。数据不仅与半保留模型的预测定量吻合,更重要的是,它以排除法的方式,干净利落地同时证伪了另外两个竞争模型。实验的严密性还体现在多个细节上:他们通过测量条带位置,计算出杂交DNA的密度增加值正好是理论预期的一半;他们通过热变性实验证明杂交分子在加热解链后,确实能分离出重链和轻链,为“一条旧链一条新链”提供了进一步证据。
这个实验没有复杂的统计学,没有晦涩的数学模型,其结论的力度直接来自于实验设计与自然现象之间那清晰、直观、无可辩驳的对应关系。这正是其被誉为“最优雅”的原因——用最简单、最直接的方式,回答了最根本的问题。
4. 涟漪与浪潮——实验的即时反响与对分子生物学的塑形
Meselson-Stahl实验的论文一经发表,立即在分子生物学界引起了轰动。它的影响是立竿见影且多层次的。
4.1 解决核心争议,巩固双螺旋范式
实验的首要贡献,便是一劳永逸地解决了DNA复制机制的争论。它提供了确凿的实验证据,支持沃森和克里克基于双螺旋结构提出的半保留复制假说。这不仅仅是验证了一个细节,而是巩固了整个分子生物学的核心范式。它证明,DNA不仅结构上是双螺旋,其功能机制也完全内嵌于这种结构之中。遗传信息的传递,本质上就是互补链的合成。从此,半保留复制从假说变为公认的事实,写入所有生物学教科书。
4.2 方法论上的革命:树立“美丽实验”的标杆
该实验在方法论上影响深远。它展示了如何将物理化学的精密手段与清晰的生物学问题相结合,创造出决定性的结果。它成为了假设-演绎法在生物学中应用的典范。科学家们意识到,解决复杂的生物学问题,不一定需要极其复杂的技术,但一定需要极其清晰的思想和严谨的设计。它激励了一代又一代的研究者去追求实验的“优雅”和逻辑的纯粹性。
4.3 技术遗产:CsCl密度梯度离心的普及
实验的成功,使得CsCl平衡密度梯度离心技术一举成为分子生物学实验室的常规利器。在接下来的二三十年里,这项技术被广泛应用于:
- 分离不同密度的核酸:如分离卫星DNA、区分不同GC含量的DNA、分离RNA-DNA杂交分子等。
- 纯化质粒DNA:在溴化乙锭染色结合CsCl梯度离心法被开发出来之前,这是纯化质粒的主要方法。
- 研究DNA复制与修复:通过类似的重轻标记策略,研究复制起点、方向、复制叉动力学以及DNA损伤修复的机制。
这项技术本身,就是Meselson-Stahl实验留给后世的一份宝贵工具遗产。
4.4 开启新的研究领域
实验的结论并没有终结研究,反而开启了一系列更深入的问题:
1.复制机制的具体细节:既然复制是半保留的,那么细胞是如何精确地解开双螺旋、合成新链、并保证高度保真度的?这直接引导了对DNA解旋酶、DNA聚合酶、单链结合蛋白、引物酶、连接酶等一系列复制机器蛋白的发现和研究。
2.复制起点与方向:DNA分子从何处开始复制?是单向还是双向?Meselson后来与其他人合作,利用类似的密度标记结合遗传学方法,在大肠杆菌和噬菌体中证明了DNA复制是双向的,并且有固定的复制起点。
3.染色体复制与细胞周期:DNA的半保留复制如何与整个细胞周期协调?如何确保每条染色体只复制一次?这连接起了DNA复制与细胞周期调控的广阔领域。
可以说,Meselson-Stahl实验像一把钥匙,打开了通往现代分子遗传学核心地带的大门,为后续数十年的爆炸性发现铺平了道路,推动了整个分子生物学学科的迅猛发展。
5. 超越时空的启示——实验的哲学意蕴与现代回响
时至今日,在基因组学、表观遗传学、合成生物学飞速发展的时代,回顾Meselson-Stahl实验,我们依然能从中汲取深刻的启示。
5.1 科学之美:简洁、清晰与必然性
这个实验常被置于“科学之美”的讨论中心。它的美体现在多个维度:
- 逻辑之美:从问题到假设,到可区分的预测,再到实验验证,线条清晰流畅,如几何证明般严谨。
- 简洁之美:核心思想非常简单——用重量标记亲代,用密度区分子代。
- 必然性之美:实验结果一旦出现,其结论似乎具有一种“必然如此”的强迫性。自然规律通过精心设计的实验,自己彰显了出来。
这种美不是装饰性的,而是认知性的。它让复杂的真理以一种易于理解和信服的方式呈现,极大地增强了科学知识的传播力和说服力。
5.2 跨学科融合的典范
实验是物理学、化学和生物学思想与技术深度融合的产物。梅塞尔森的物理化学背景与斯塔尔的遗传学背景缺一不可。这预示了现代生命科学发展的根本路径——日益依赖并深度融合数理科学、工程学和信息科学的技术与思维。从结构生物学到系统生物学,再到今天的计算生物学和AI驱动的新药发现,跨学科融合已成为创新的主要引擎。
5.3 对现代研究的隐喻与延续
Meselson-Stahl实验的精神在现代研究中以新的形式延续:
- “重轻”标记的现代变体:虽然CsCl离心已较少使用,但同位素标记和密度/重量区分的思想仍然是研究生物大分子合成、降解、修饰和相互作用的基石。
- 单分子水平的“复制”观测:现代单分子技术允许科学家实时观测单个DNA分子被解旋、复制的动态过程,这可视作对Meselson-Stahl群体平均结果的终极单分子验证和深化。
- 表观遗传信息的“复制”问题:今天我们知道,遗传不仅关乎DNA序列,还关乎其上的化学修饰。
这些“表观遗传”信息在细胞分裂中如何传递?是“全保留”、“半保留”还是“分散”模式?这成为了一个新兴的热点领域,其研究范式常常回溯和借鉴Meselson-Stahl实验的思想。
5.4 科学文化与教育意义
该实验是科学教育的绝佳案例。它教导学生:
- 如何提出一个好问题:问题要根本、要清晰。
- 如何设计判决性实验:关键在于推导出竞争理论的不同预测。
- 如何解读数据:数据要直接、可视化,结论要源于数据与预测的严格比对。
- 欣赏科学过程的创造性:科学发现不仅是积累数据,更是智慧、想象力和严谨性的结晶。
结论:一座永恒的灯塔
自1958年那个决定性的实验完成以来,六十多年过去了。分子生物学已经从描述双螺旋结构,发展到可以编辑基因组、合成人工生命。然而,Meselson-Stahl实验的地位丝毫未曾动摇。它不仅仅是一个历史事件,更是科学精神的一座灯塔。
它照亮了从分子结构到生命功能的认知之路,证明了基于深刻理论洞察的、设计精良的实验,拥有改变整个学科面貌的力量。它体现了科学中理性与美感的高度统一,展示了人类智慧如何通过巧妙的工具与自然对话,并解读出生命最基本奥秘之一的答案——遗传的蓝图,正是通过那优雅的半保留方式,在世代间忠实地传递。
在当今这个数据爆炸、技术纷繁复杂的时代,重温Meselson-Stahl实验,提醒我们:无论技术如何进步,清晰的科学思维、严谨的逻辑演绎和追求本质的简单性,永远是科学发现最宝贵的品质。这份遗产,将与其证明的DNA半保留复制机制一样,在科学的长河中,被永久地传承下去。


近期评论