

 扫码安装网站APP(Android版)
扫码安装网站APP(Android版)
来源公众号:鹤风细语
【引言】大约在3.5亿年前大气中CO₂含量明显下降而O₂浓度显著增加,导致叶绿体的光呼吸增强,这种变化驱使植物进化出一系列适应机制,积极地从环境吸收CO₂和HCO₃⁻,达到在Rubisco周围积累无机碳的目的,进而减少加氧反应,降低光呼吸。
本文介绍在Rubisco羧化位点进行CO₂浓缩的一种常见机制:C4光合作用的碳固定———C4循环。
一.C4循环的发现
20世纪50年代晚期,H.P.Kortschack和Y.Karpilov分别利用¹⁴CO₂标记的甘蔗和玉米进行实验。在光下,接受¹⁴CO₂处理几秒后,便发现叶片中70%~80%的¹⁴C出现在苹果酸和天冬氨酸等四碳酸中,这种现象在只能进行卡尔文循环的叶片中没有发生。
1966年,M.D.Hatch和C.R.Slack提出了四碳二羧酸途径,简称C4循环,或C4途径。具有这种碳同化途径的植物称为C4植物。这类植物大多起源于热带或亚热带,主要集中于禾本科、莎草科、菊科和苋科等科。
二.C4循环的结构基础
研究发现,进行C4循环的植物,其叶片有特殊的结构特征,如下图所示。
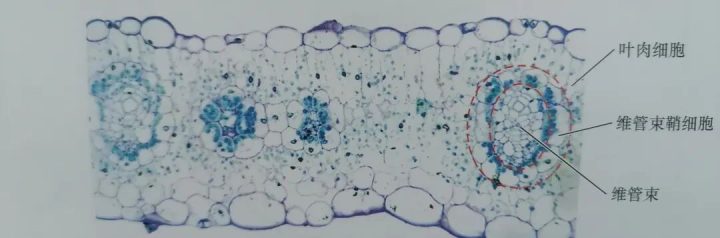
1. 花环状结构
维管束鞘细胞发达:C4植物的维管束鞘细胞体积大、壁厚、含有大量叶绿体,且叶绿体通常无基粒或基粒不发达(适应碳浓缩反应)。叶肉细胞环绕排列:叶肉细胞围绕维管束鞘细胞呈放射状排列,形成类似“花环”的结构,这种紧密排列缩短了细胞间物质运输距离。
2. 叶绿体分化(两种类型)
叶肉细胞叶绿体:主要负责进行C4途径的初步固碳(由PEP羧化酶催化固定CO₂,形成四碳化合物),同时还为维管束鞘细胞光合作用提供ATP和NADPH。
维管束鞘细胞叶绿体:负责进行卡尔文循环,将C4化合物释放的CO₂进一步合成糖类。其叶绿体常具有淀粉积累能力。
3. 胞间连丝密集
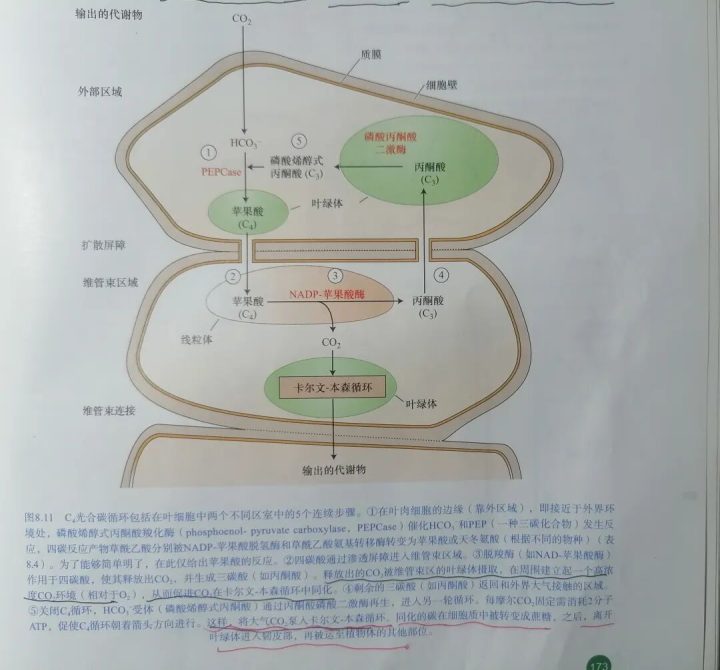
叶肉细胞与维管束鞘细胞之间胞间连丝数量多,便于C4化合物(如苹果酸、天冬氨酸)快速转移,以及三碳化合物丙酮酸或丙氨酸经由胞间连丝运回叶肉细胞。三.C4循环的反应过程: CO₂从外部空气中到被运输至维管束鞘细胞内参与卡尔文循环,经历了5个步骤。
1.羧化(在叶肉细胞进行):C4循环的CO₂受体是叶肉细胞质中的PEP(磷酸烯醇式丙酮酸),在磷酸烯醇式丙酮酸羧激酶的催化下,固定HCO₃⁻,生成草酰乙酸。随后,草酰乙酸被NDP-苹果酸脱氢酶还原成苹果酸,或者和谷氨酸进行转氨基作用产生天冬氨酸。
2.转运(通过胞间连丝):四碳酸(苹果酸或天冬氨酸)通过胞间连丝转运到维管束鞘细胞。
3.脱羧(在维管束鞘细胞):四碳酸(苹果酸或天冬氨酸)脱羧变成丙酮酸或丙氨酸,释放的CO₂进入卡尔文循环还原成糖类物质。
4.返运(通过胞间连丝):由脱羧反应产生的三碳化合物丙酮酸或丙氨酸经由胞间连丝运回叶肉细胞。
5.再生(在叶肉细胞):丙酮酸或丙氨酸在叶肉细胞叶绿体中,在丙酮酸磷酸二激酶催化和ATP作用下,生成CO₂受体PEP,使反应持续进行。
这5个步骤的反应体现在下图中:
四.维管束鞘细胞光合作用所需ATP和NADPH的来源
在C₄植物中,维管束鞘细胞是进行卡尔文循环(碳同化)的主要场所。然而,维管束鞘细胞通常缺乏完整的PSII系统(光系统II),导致其光反应能力有限,无法完全独立产生足够的ATP和NADPH。因此,维管束鞘细胞光合作用所需的ATP和NADPH主要通过以下两种方式获得:
1. 通过胞间连丝从叶肉细胞输入。叶肉细胞进行完整的光反应,生成大量ATP和NADPH。部分ATP和NADPH(或以其还原等价物形式)通过胞间连丝直接转运到维管束鞘细胞,用于卡尔文循环。
2. 维管束鞘细胞自身的光反应贡献。 部分C₄植物的维管束鞘细胞仍保留一定光反应能力。主要依赖光系统I(PSI)进行环式光合磷酸化,产生少量ATP(不产生NADPH和O₂)。 因无法通过非环式光合磷酸化生成NADPH(因缺乏PSII), 所以,NADPH主要依赖叶肉细胞。
结论:维管束鞘细胞光合作用所需的ATP和NADPH主要依赖叶肉细胞通过胞间连丝供应,自身光反应仅能有限提供ATP,这是C₄植物叶肉细胞和维管束鞘细胞进行分工合作,优化光合策略的重要特征。
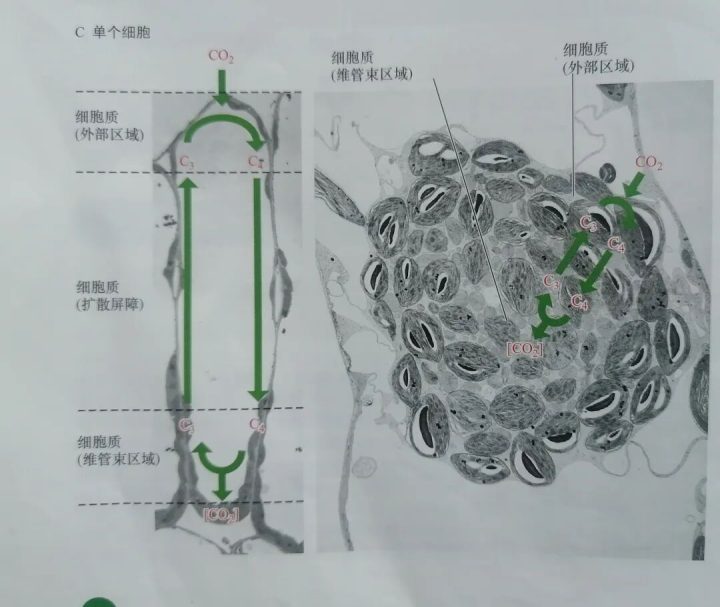
五.单个细胞中的C4循环也能够浓缩CO₂
研究发现,有些植物的C4光合作用不具有“花环”结构,它们能够在单个细胞中完成整个C4光合作用。基本过程如下图所示:
六.C4循环的生理学意义
植物C4循环是一种高效的光合作用途径,在生理学上具有重要的适应意义,主要体现在以下几个方面:
1. 减少光呼吸,提高光合效率
在高温、强光、干旱条件下,植物气孔关闭导致叶片内CO₂浓度降低,而O₂浓度相对升高。Rubisco酶(参与卡尔文循环的关键酶)在O₂浓度高时会发生光呼吸,消耗能量并释放CO₂,降低光合效率。
C4植物通过空间分离(在叶肉细胞和维管束鞘细胞中分别进行)将CO₂的固定与还原过程分开。首先在叶肉细胞中,PEP羧化酶(对CO₂亲和力极高)固定CO₂生成四碳化合物(如草酰乙酸),随后将CO₂“泵”入维管束鞘细胞释放,使Rubisco周围维持高浓度CO₂,从而抑制光呼吸,提升光合效率。
2. 节约水分,减少氮素利用
节水机制:PEP羧化酶对CO₂的高亲和力允许C4植物在气孔部分关闭时仍能有效固定CO₂,从而减少蒸腾作用的水分流失,水分利用效率(单位水消耗固定的CO₂)可比C3植物高1.5–3倍。
氮素利用高效:Rubisco酶含量在C4植物中较低(因CO₂浓缩机制降低其需求),而Rubisco是植物叶片中含量最多的蛋白质,合成需消耗大量氮素。因此C4植物单位氮素的光合效率更高,适应贫瘠土壤。
4. 在农业方面的应用:
C4作物通常具有更高的生物产量潜力,尤其在高温强光条件下(如玉米、甘蔗的高产特性)。研究C4机制可为作物改良(如将C4特性导入C3作物)提供方向。目前科学家正尝试通过基因工程将C4光合机制引入水稻等C3作物,以提高产量和抗逆性。这一方向展示了C4生理机制在应对气候变化和粮食安全挑战方面的潜在价值。
【综述】:C4循环是植物在长期进化中形成的高效适应策略,通过生化与结构的协同优化,解决了光呼吸的能耗问题,从而在高温、干旱、强光及低CO₂条件下维持较高的光合生产力。将来,通过基因工程手段把C4机制引入C3作物,为保障人类粮食安全具有十分诱人的潜在价值。
来源网址:高中生物拓展知识:C4植物的光合作用



近期评论