

 扫码安装网站APP(Android版)
扫码安装网站APP(Android版)
来源公众号:通俗生物学 作者:Ffm18234
摘要:叶绿体作为植物和藻类进行光合作用的关键细胞器,其起源与内共生事件密切相关。本文系统综述了叶绿体的内共生起源学说,详细探讨了其从原始蓝细菌演化为主营光合作用的细胞器的过程,并深入分析了叶绿体在生物进化中的关键作用。研究表明,叶绿体的出现不仅推动了光合自养生命的繁荣,促进了地球氧化环境的形成,还通过基因转移和协同进化深刻影响了真核生物的演化路径。本文还结合最新研究进展,探讨了叶绿体进化研究中的前沿问题与争议,旨在为理解生命与环境互作提供更全面的视角。
1. 引言
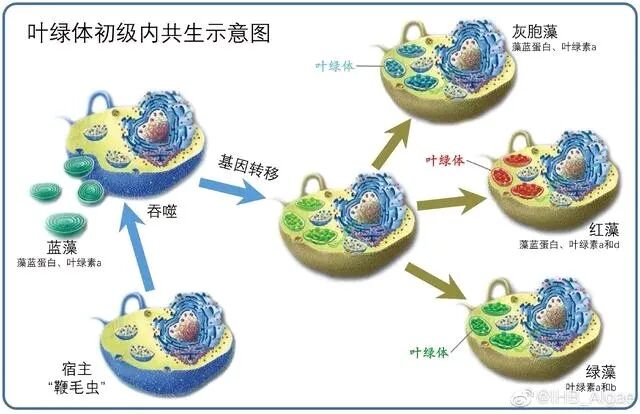
叶绿体是光合真核生物的能量转换中心,能够利用光能将二氧化碳和水转化为有机物,并释放氧气。现代生物学普遍认为,叶绿体起源于约10亿年前的一次原始真核细胞吞噬蓝细菌并形成稳定共生关系的事件,这一过程被称为“初级内共生”。此后,叶绿体通过二次或多次内共生事件在不同藻类及植物中扩散,形成了多样化的光合生物类群。叶绿体的出现不仅改变了地球的碳氧循环,也对生物进化轨迹产生了深远影响。随着基因组学、结构生物学和合成生物学的发展,叶绿体起源与进化的研究不断深入,揭示了更多关于细胞器形成、基因转移机制以及宿主-共生体互作的奥秘。
2. 叶绿体的起源:内共生学说
2.1 内共生学说的提出与历史背景
内共生学说(Endosymbiotic Theory)最早由康斯坦丁·梅列施柯夫斯基(Konstantin Mereschkowsky)于1905年提出,他认为叶绿体是由原先的内共生体形成的。随后,琳·马古利斯(Lynn Margulis)在20世纪60年代系统阐述了这一理论,并指出线粒体和叶绿体分别起源于好氧细菌和蓝细菌的内共生事件。尽管最初受到质疑,但随着分子生物学证据的积累,内共生学说已成为解释真核细胞器起源的主流理论。
2.2 内共生学说的详细证据
内共生学说得到了多方面的实验支持,其证据主要包括以下几个方面:
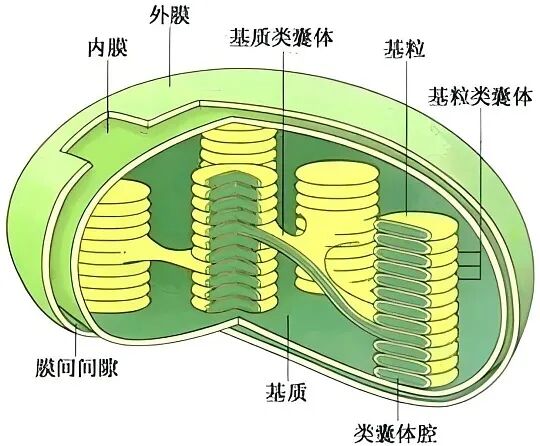
(1)形态与结构相似性:叶绿体与蓝细菌在大小、膜结构(特别是类囊体)和光合色素组成上高度相似。叶绿体被双层膜包被,其中最里面一层的成分与细胞中其它膜不同,更接近原核生物的细胞膜。此外,叶绿体内部的类囊体结构与蓝细菌的类囊体排列方式相似,且都含有叶绿素a和类胡萝卜素等光合色素。
(2)遗传独立性:叶绿体拥有自身的环状DNA、核糖体及转录翻译系统,与细菌遗传体系相近。叶绿体的核糖体由30S和50S两个亚基组成,与真核生物细胞质中的核糖体(40S和60S)不同,而与蓝细菌的核糖体一致。抗生素(如氯霉素)可以抑制细菌和蓝藻的生长,也可以抑制真核生物中叶绿体的作用,这说明叶绿体与蓝藻是同源的。
(3)系统发育分析:叶绿体基因的分子系统学研究表明,其与现存蓝细菌的亲缘关系密切。例如,蓝藻的核糖体RNA(rRNA)不仅可以与蓝藻本身的DNA杂交,还可与眼虫叶绿体的DNA杂交,这些都说明它们之间的同源性。DNA序列分析显示,核DNA包含了一些可能来源于叶绿体的基因片段,支持了基因转移的假说。
(4)内共生过程的模拟与观察:实验显示,某些原生生物可摄入藻类并建立临时共生,为长期内共生提供可能途径。例如,灰孢藻(Glancocystis)体内有叶蓝小体(cyanella)进行光合作用,这种共生关系建立不久,叶蓝小体在细胞内还不固定,是对内共生假说的有力支持。
2.3 叶绿体的进化阶段
叶绿体的进化经历了吞噬、基因转移和代谢整合等多个阶段:
(1)吞噬与共存:原始真核细胞吞噬好氧蓝细菌,但未将其消化,而是形成共生关系。蓝细菌在宿主细胞内存活下来,并逐渐演化为叶绿体。
(2)基因转移与精简:蓝细菌的大部分基因转移至宿主细胞核,叶绿体基因组大幅缩减,形成依赖宿主调控的半自主细胞器。例如,原始蓝细菌的基因组约为3 Mb,而现存叶绿体的基因组通常小于200 kb,超过90%的基因通过内共生基因转移(EGT)转移至核基因组。
(3)代谢整合:宿主与内共生体之间形成代谢物交换机制,如宿主通过质膜转运蛋白(如NTT1)向叶绿体输送ATP,叶绿体则通过光合作用提供有机物和能量。
3. 叶绿体在生物进化中的关键作用
3.1 推动光合自养生物的辐射演化
叶绿体的形成使真核生物获得了光合自养能力,直接促进了绿色植物、红藻、褐藻等多样光合类群的分化与繁盛。这些光合生物构成了陆地和水生生态系统的初级生产力基础,支撑了食物链的能量流动。例如,绿藻和红藻的叶绿体显示出不同的结构和功能,表明它们的演化历程并不一致,可能通过多次内共生事件获得各自独特的叶绿体。
3.2 改变地球环境与生物圈
(1)大氧化事件(GOE)的延续:叶绿体的大规模光合产氧作用进一步提升了大气氧含量,促进了需氧真核生物的演化及臭氧层的形成。氧气浓度的增加为复杂多细胞生物的出现提供了必要条件。
(2)碳循环的重构:叶绿体驱动的光合作用加速了碳的生物固定,减少了大气中的二氧化碳浓度,影响了全球碳循环与气候。光合作用产生的有机物为生物圈提供了能量来源,推动了生态系统的复杂化。
3.3 促进真核生物的基因与代谢进化
(1)水平基因转移(HGT):叶绿体基因向核基因组的转移丰富了宿主遗传信息,促进了核基因组的复杂性提升。研究表明,细胞器DNA向核基因组的转移(NORG)在植物演化过程中持续发生,每百万年约有631–944个插入事件,这些转移增加了核基因组的遗传多样性。
(2)代谢网络整合:叶绿体与线粒体、过氧化物酶体等细胞器协同,形成了高效的能量与物质代谢网络。叶绿体不仅参与光合作用,还参与合成脂肪酸、氨基酸以及植物激素前体,直接支持细胞代谢与发育。
3.4 为生物内共生理论提供范式
叶绿体的内共生起源是“细胞器通过内共生形成”的最经典例证,为理解线粒体等细胞器的起源提供了理论基础。内共生学说不仅解释了细胞器的起源,还揭示了生物进化中的协作与互惠关系,挑战了传统的竞争进化观。
4. 叶绿体基因转移与进化机制
4.1 基因转移的机制与规律
叶绿体基因向核基因组的转移主要通过DNA介导的机制实现,而非通过mRNA/cDNA中间体。研究表明,细胞器DNA的转移主要通过双链断裂修复途径(如非同源末端连接c-NHEJ)和复制依赖的机制(如FoSTeS、MMBIR)形成。转座元件在造成NORG片段化的演化过程中也发挥了重要作用。
4.2 基因丢失与补偿机制
叶绿体基因组在进化过程中经历了显著的基因丢失,例如编码翻译起始因子1的infA基因在多数被子植物中已丢失,但通过核基因组的功能性拷贝得以补偿。寄生植物中的叶绿体基因丢失尤为显著,如全寄生植物多室花科(Lennoaceae)中,近半数质体基因丢失或假基因化,仅保留管家基因。
4.3 结构重排与进化多样性
叶绿体基因组通常结构稳定,但某些物种中仍存在显著的结构重排。例如,反向重复区(IR)的边界迁移、扩张与收缩是驱动质体基因组大小变异和结构多样性的关键机制。重复序列介导的重组事件可能导致基因重复或丢失,进一步增加了叶绿体基因组的多样性。
5. 争议与前沿问题
5.1 内共生发生的具体次数与途径
关于叶绿体的起源是单次还是多次内共生产物仍存争议。早期研究认为所有叶绿体都源于一次共生事件,但近年来的研究表明,不同生物群体中的叶绿体可能来自多次内共生事件。例如,红藻和绿藻的叶绿体显示出不同的特征,支持多重内共生的假说。
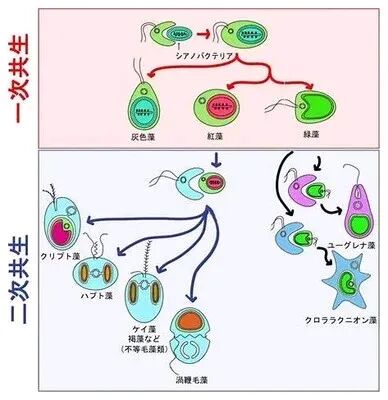
5.2 基因转移的驱动机制
核与叶绿体间基因转移的选择优势与调控机制尚未完全阐明。尽管研究表明基因转移在进化中持续发生,但转移后的基因如何整合到核基因组并发挥功能仍需进一步研究。例如,转移的基因如何获得启动子及正确的RNA加工信号,以及如何避免有害突变的影响。
5.3 叶绿体蛋白转运机制的进化
叶绿体蛋白转运系统(如TOC-TIC复合体)的进化机制是当前研究的热点。最新研究表明,Ycf2-FtsHi复合体是叶绿体蛋白转运的动力源(马达),其在绿色植物谱系中的进化多样性反映了叶绿体与宿主细胞的协同演化。例如,拟南芥和莱茵衣藻中的TOC-TIC复合体结构存在差异,揭示了进化过程中的保守性和多样性。
5.4 合成生物学应用
人工构建“合成叶绿体”或改造光合系统,可能为农业和能源领域带来突破。例如,通过调控叶绿体门控系统的效率,优化光合作用效率,提高作物产量。实验室尝试将蓝藻导入斑马鱼胚胎细胞或酵母,通过基因编辑建立临时共生,为理解内共生建立的分子壁垒提供了新视角。
6. 结论
叶绿体的出现是生命进化史上的一次革命性事件。通过内共生起源,叶绿体不仅赋予了真核生物光合能力,重塑了地球生态环境,还通过基因和代谢整合深刻影响了真核细胞的进化方向。未来研究需结合比较基因组学、细胞进化学与合成生物学,进一步揭示叶绿体进化细节及其在生命系统中的作用。随着技术的进步,我们对叶绿体起源与进化的理解将不断深化,为生物进化理论提供更多支持。
来源网址:叶绿体的起源及其在生物进化中的作用



近期评论