


 扫码安装网站APP(Android版)
扫码安装网站APP(Android版)
来源公众号:地球玩家2号
高中生物老师备课参考
以图文形式,系统讲解二次应答更快更强”的分子机制;从抗体结构入手,把”免疫记忆”从现象层挖到分子层,给老师一份”心里有底”的备课参考。
开篇:一个学生答得出、老师答不全的问题
先看一道几乎所有高三生都做过的题目——关于”二次免疫应答比初次更快、更强”的判断。学生都会做。但如果你追问一句:”为什么更强?”——能从分子层面答上来的,凤毛麟角。
翻开课本和其他教辅,关于免疫记忆的描述甚少,对于为什么二次产生的抗体不仅多,而且”更管用”,也无系统而全面解释。
这些问题不是钻牛角尖。它们是学生理解免疫这一章的关键节点,也是老师备课时最容易”心虚”的地方。本文要做的,就是把背后的分子机制讲透——既不超出”高中加深拓展”的合理边界,又能让你面对学生提问时给出有底气的回答,而不是一句”以后大学会学”。
先看一张图。这张图你一定在教辅上见过
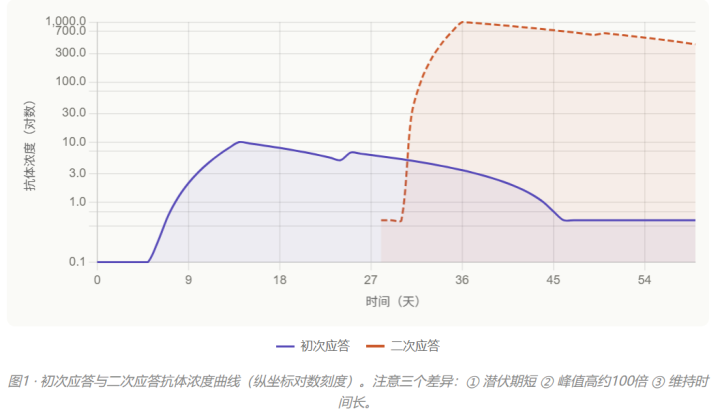
记住这三个特征:①二次免疫的潜伏期更短(潜伏期为1~2天)、②峰值高(比初次应答高约100倍)、③维持时间更长。
文会一一回答它们各自来自哪里
一、先破一个误区:”记忆细胞”不是一种细胞
教材的简化与代价
高中教材把”记忆细胞”作为一个统一概念引入,简洁但模糊。学生最常见的误解是:以为”记忆细胞=能产生抗体的细胞”。这是错的。记忆B细胞本身不分泌抗体——它要在再次遇到抗原后,分化为浆细胞,才开始分泌。
更深一层的事实是:免疫记忆从来不是某一种细胞的事,而是至少三类细胞各司其职的结果。

图2 · 三类记忆细胞分工。讲”二次应答”时模糊处理为”记忆细胞”可行,但学生追问时心里要有这三个分工。
二、第一层机制:数量优势
2.1 最容易理解的一层
这是大多数教辅会讲到的一层——也是最浅的一层。初次免疫应答时,能够识别这种抗原的B细胞和T细胞会被大量”克隆扩增”。应答结束后,绝大多数效应细胞(如浆细胞、效应T细胞)会凋亡,但约5%到10%的细胞会分化为记忆细胞保留下来。
这些保留下来的记忆细胞,数量远高于初次应答前的初始淋巴细胞。来感受一下量级:
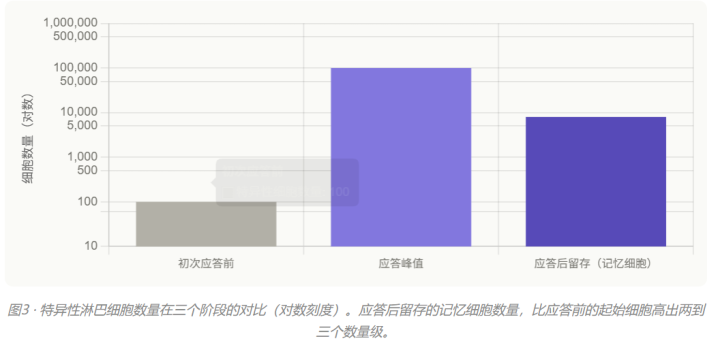
所以当病原体第二次入侵时,能够识别它的细胞数量已经从”几十到几百”变成了”成千上万”。这是二次应答”启动更快”的第一个原因:起跑线本身就高了两到三个数量级。
2.2 但这一层不够
仅靠数量优势,无法解释一个关键事实——二次应答产生的抗体,质量也更高。同样是10万个浆细胞,二次应答产生的抗体与抗原的结合能力,比初次应答的抗体强几十甚至上千倍。
这就引出了下一层、也是教材完全没讲的核心机制——质量优势。但在讲清楚”质量”之前,我们必须先把”抗体”这个分子的结构搞清楚——这是后面所有讨论的基础,也是不少老师备课时容易模糊的一块。
三、必备基础:抗体到底长什么样?
要理解后文的”亲和力成熟”和”类别转换”,我们必须把抗体分子拆开看。
3.1 一个抗体 = 四条肽链
一个抗体分子由四条多肽链组成:
- 2 条相同的重链(H 链),每条约 450 个氨基酸;
- 2 条相同的轻链(L 链),每条约 215 个氨基酸。
四条链通过二硫键相互连接——重链与重链之间、每条重链与一条轻链之间,都有二硫键交联。最终组合成一个稳定的 Y 字形 结构
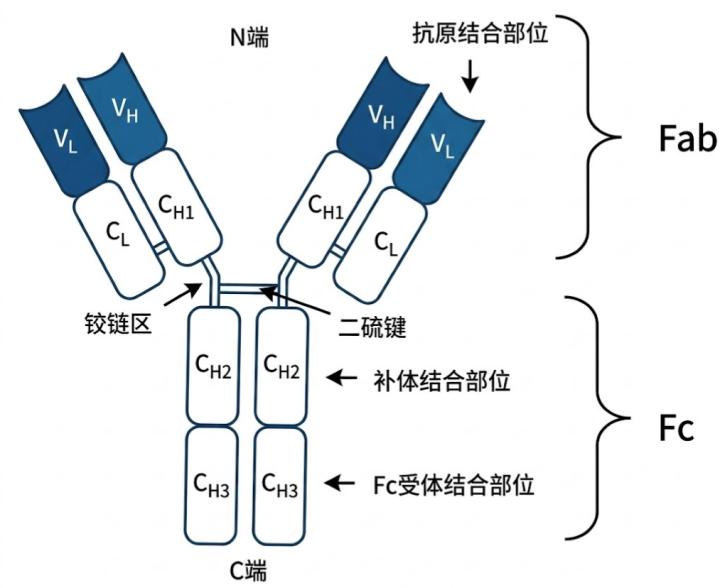
图4 · 抗体分子结构详解。Y形分子由2条重链(H)和2条轻链(L)通过二硫键连接而成;每条链都分为可变区(V)和恒定区(C);Y的两个”臂”叫Fab,”柄”叫Fc。
3.2 可变区 vs 恒定区
四条链上每个区段,又各自分为可变区(V,Variable)和恒定区(C,Constant)。命名是字面意思——
- 可变区:氨基酸序列在不同抗体之间差异极大。它位于Y的两个臂的顶端,是真正接触抗原的部位。
- 恒定区:氨基酸序列在同一类抗体中高度保守。它构成抗体的”骨架”和”柄部”,决定抗体的效应功能。
3.3 Fab 和 Fc:功能上的两个区域
如果我们按功能把抗体切成几块,就得到两个关键区域:
Fab 区域:Y 字的两个”臂”。每个 Fab 由 整条轻链 + 重链的 VH 和 CH1 段 组成。Fab 顶端的 VH+VL 联合形成的口袋,就是抗原结合位点。一个抗体有两个 Fab,所以能同时结合两个抗原表位。
Fc 区域:Y 字的”柄”。由 两条重链的 CH2 + CH3 段 组成(IgM/IgE还有CH4)。Fc 不结合抗原,而是结合免疫细胞表面的 Fc 受体——激活补体、招募吞噬细胞、被NK细胞识别等。Fc 的氨基酸序列决定了抗体是 IgM 还是 IgG,决定了抗体的”派别”。
中间还有一段叫铰链区(hinge region),位于 CH1 和 CH2 之间,富含脯氨酸,结构柔韧,让两个 Fab 臂可以灵活张合,适应不同间距的抗原表位。
3.4 一句话总结这个结构
可变区(V)= 在Fab尖端 = 决定”识别什么”
恒定区(C,尤其是Fc段)= 在Y的柄部 = 决定”是哪类抗体、做什么”
这个对应关系,是后面理解亲和力成熟和类别转换的钥匙。
有了这个结构基础,下面要讲的两个机制就一目了然了——“亲和力成熟”改的是Fab尖端的V区;”类别转换”改的是Y柄部的C区。两件事各自只动抗体的一部分,互不干扰。这种”模块化”恰恰是生命系统最精彩的工程设计之一。
四、第二层机制(核心难点):亲和力成熟
先问自己一个问题
二次应答产生的抗体,和初次应答的抗体是同一种抗体吗?
直觉答案是:”是啊,都是针对同一种抗原,蛋白质结构应该一样。”
真实答案是:不是同一种抗体——它们的氨基酸序列不同,与抗原的结合亲和力差几十到上千倍。
这就是免疫学里一个被高中教材完全省略、但又极其精彩的概念——亲和力成熟。
4.1 亲和力是什么?
简单说,抗体和抗原的结合就像锁和钥匙——锁齿越吻合,结合越牢固,亲和力越高。注意:这里发生变化的只是Fab尖端的VH+VL区,抗体其余部分都不变。

图5 · 抗体亲和力的视觉对比。同一个抗原表位,左边是初次应答抗体的Fab尖端,结合面有缝隙;右边是经过突变筛选后的二次应答抗体,结合面紧密贴合。变化只发生在VH+VL两段。
4.2 亲和力成熟靠什么实现?——体细胞高频突变
B细胞中存在一种叫 AID(活化诱导胞苷脱氨酶)的酶,它的作用是——专门针对编码VH和VL(即抗体可变区)的基因片段,引入高频点突变。这个过程称为体细胞高频突变。
这一层有多”高频”?
体细胞高频突变的频率比基因组其他部位的常规突变率高约10⁶倍。换句话说,B细胞主动在抗体识别区域制造突变——这是一种”有目的的突变”。
但请注意:突变本身是随机的,筛选才是非随机的。这是理解整个机制的关键。
4.3 生发中心:微观尺度的”突变-筛选”
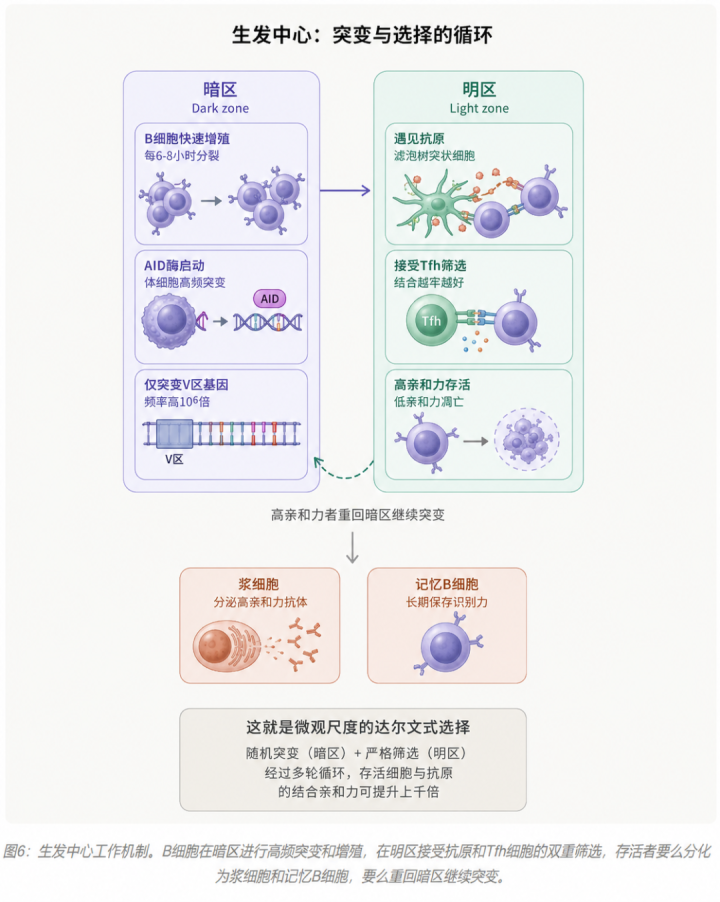
生发中心由两个功能区组成:
- 暗区(Dark zone):B细胞密集增殖,每6到8小时分裂一次。AID酶在这里启动,对V区基因进行高频突变。
- 明区(Light zone):突变后的B细胞带着新版本的”抗体钥匙”,去测试能否与抗原结合得更牢。结合越牢的细胞,越能获得Tfh细胞(滤泡辅助性T细胞)的存活信号;结合不牢的,凋亡。
关键在于:存活下来的高亲和力B细胞,会重新回到暗区,再来一轮突变和筛选。如此循环若干次。这就是一个微观尺度的、几天之内完成的“突变-选择”循环。
4.4 与进化论的精彩呼应
这里有一个跨单元的关联值得指出:高中教材在”现代生物进化理论”中讲过”突变是进化的原材料,自然选择决定方向”。但学生很难想象——这种”突变-选择”在个体一生中、几天之内就能发生。
亲和力成熟正是这样一个例子。进化论的逻辑,正在每个人的淋巴结里每天上演。
回到前面提出的问题:”记忆细胞再次遇到抗原时产生的抗体与初次相同吗”,严格意义上,氨基酸序列不完全相同,亲和力差几十到上千倍——这是给优秀学生留的口子。
五、第二层机制(续):类别转换
5.1 又一个被教材回避的真相
翻开教材,关于抗体的描述基本停留在”由B细胞分泌的免疫球蛋白”。但实际上,抗体并不是一种均一的分子,而是分为五大类——IgM、IgG、IgA、IgE、IgD。它们的差别就在恒定区,准确说是重链的恒定区。
这里有一个老师备课时应该知道的关键事实:初次应答 vs 二次应答的抗体类别差异
初次应答主要产生 IgM;二次应答主要产生 IgG(以及 IgA、IgE 等)。
两者结构、寿命、功能都不同。这种切换叫抗体类别转换。
5.2 IgM和IgG的差别为什么重要?
- IgM:由5个Y形单元组合成五聚体(重链恒定区是Cμ,多了一个CH4结构域),分子很大,主要在血液中循环;激活补体能力强,但组织穿透性差。半衰期约5天。
- IgG:单体(重链恒定区是Cγ),分子小,能穿过胎盘(这就是为什么新生儿能从母亲那里获得短期免疫);半衰期约21天,是长期保护的主力。
用一个比喻:IgM 是急救兵——一旦感染立即上场,火力猛,但持续时间短;IgG 是常备军——反应稍慢但持久作战,是免疫记忆的长期产物。
5.3 类别转换的分子本质
有了前面打下的结构基础,这件事就好讲了:
关键事实:重链基因的可变区(V)和恒定区(C)在DNA上是分开排列的——一段V基因后面,紧跟着 Cμ、Cγ、Cα、Cε 等多个恒定区基因段,依次排列。
最初的B细胞默认表达”V + Cμ”,所以分泌的是IgM。类别转换的本质,就是把不需要的Cμ段切除,让V直接连接到Cγ(或Cα、Cε)。
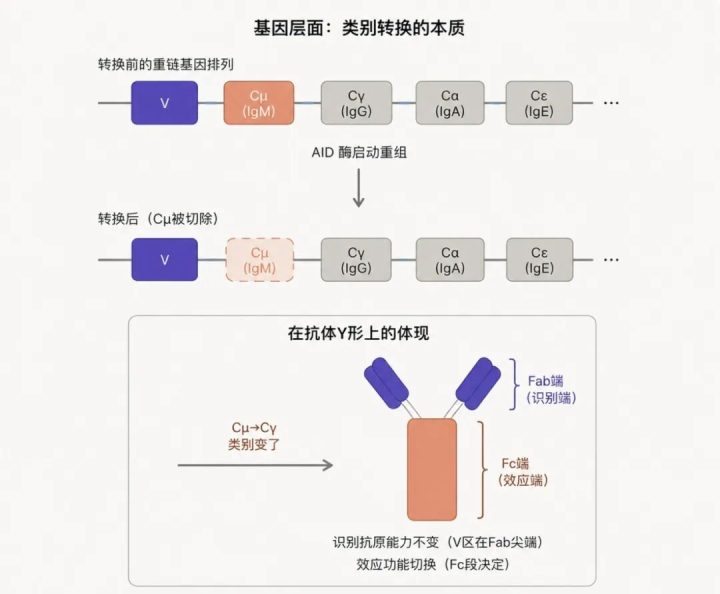
图7 · 抗体重链基因的类别转换。AID酶启动DNA层面的重组,把Cμ基因段切除,让V直接连接到Cγ。在抗体Y形结构上的体现是:Fab尖端的V区不变(识别能力保留),Fc段从Cμ变成Cγ(功能改变)。
这个过程同样由AID酶启动(没错,亲和力成熟和类别转换共用一个核心酶——这也是AID酶被发现时震动免疫学界的原因之一)。同一个B细胞,在生发中心可以经历一次或多次类别转换。
5.4 这件事对学生理解”基因-蛋白质关系”很重要
教材讲基因控制蛋白质合成时,呈现的是”一个基因 → 一种 mRNA → 一种蛋白质”的简化模型。这个模型不错,但学生很容易固化为”一对一”的强对应。
类别转换是这个模型的精彩反例:同一个B细胞,先后表达不同类别的抗体,靠的是DNA层面的重组——把不需要的基因段直接切除掉。这告诉学生:基因表达不是简单的”读取”,还可能涉及基因本身的重排。
六、第三层机制:启动优势
数量讲完了,质量讲完了,还剩最后一个谜题——为什么二次应答”反应这么快”?仅仅是因为细胞多吗?
不是。即便细胞数量相同,记忆细胞的反应速度也明显快于初始淋巴细胞。这是因为记忆细胞本身就处于一种“半激活的待命状态”。
6.1 记忆细胞的几个内在差异
- 表面受体亲和力高:记忆B细胞表面已经携带高亲和力的BCR(B细胞受体,本质是膜结合形式的抗体),而初始B细胞的BCR亲和力较低。这是亲和力成熟留下的”遗产”。
- 共刺激分子表达高:T细胞激活通常需要两个信号——抗原识别和共刺激信号。记忆T细胞表面的共刺激分子(如CD27等)表达更高,激活所需的信号阈值更低。相关详解见免疫系统 90% 的时间在干一件事:不要攻击
- 代谢半激活:记忆细胞的基础代谢率高于初始淋巴细胞,处于”随时可以爆发”的状态,不需要经历从静息到激活的完整启动过程。
- 位置分布优势:记忆细胞常驻在更利于快速反应的位置——淋巴结、脾、黏膜组织(如肠道、呼吸道黏膜)。当病原体再次入侵,它们就在前线,无需远程动员。
6.2 三层机制如何对应曲线特征?
现在回到开篇那张曲线图。三个特征,分别由不同的机制贡献:

图8· 免疫记忆三层机制总览。这是一张可以保存、可以反复回看的认知地图,是分子层面解读。
七、一个反例:为什么有些病我们”记不住”?
讲完了”为什么能记住”,反过来问一句”为什么记不住”,能让机制理解更深一层。流感、新冠、HIV——这些病我们每年都要担心,似乎免疫记忆对它们”失效”了。为什么?
7.1 失效的三种典型机制
① 抗原漂变。流感病毒表面的血凝素(HA)和神经氨酸酶(NA)蛋白持续突变。记忆细胞认识的是”旧版本”的表面蛋白,遇到新变异株时,抗体对应的”钥匙”已经不再吻合”新锁”。这就是流感疫苗每年都要重新研发的原因。
② 原始抗原效应。这是一个非常有趣的现象——当人体遇到与初次感染毒株相似但不完全相同的新毒株时,免疫系统倾向于优先动员针对原始毒株的记忆细胞,而不是产生全新的、针对新毒株的应答。这反而会让针对新毒株的反应不够充分。这个现象在流感和登革热研究中都有体现,这里不展开。
③ 免疫逃逸。HIV病毒最阴险的地方在于——它感染的主要细胞,恰好是辅助性T细胞(CD4+ T细胞)。也就是说,HIV从根本上摧毁了免疫应答的”总指挥”,让免疫记忆从源头上无法建立。这也是为什么AIDS的”AID”是获得性免疫缺陷。
7.2 这部分的教学价值
讲这些反例不是为了把免疫课变得复杂,而是让学生理解一个更深的道理:免疫记忆不是万能的。它依赖于抗原表位的稳定、依赖于免疫系统本身的完整、依赖于”突变-选择”循环能产生有效抗体。任何一个环节出问题,记忆就会失效。
这也回答了一个学生常问的生活问题——”为什么流感疫苗每年都要打一次,而麻疹疫苗打过就管一辈子?”答案就藏在”抗原是否漂变”这一点上。
八、结尾:
关于”讲到哪里”:高中教材的简化处理是合理的——课时有限、信息量有限,重点是建立”现象层”的认知框架。但作为老师,心里要有比讲台更深的底。当学生在课堂上、在课后追问”为什么”时,能用一两句通俗的话点透,而不是含糊带过。
讲台上不必把AID酶、生发中心、可变区/恒定区、类别转换这些名词都摆出来——超纲。但你可以这么回应学生:
- “抗体是个Y形分子,两个臂的尖端负责’识别’抗原,柄部决定’抗体属于哪类’。这两块功能分得很清楚。”
- “二次应答的抗体和初次的不完全一样,淋巴结里有一个’考场’,让B细胞自己变异、自己挑出’结合得更牢’的版本——但只变Y的尖端,柄部不动。”
- “二次应答的抗体类型也变了,从短期的IgM切换到了长期的IgG——这只是把柄部换了,识别能力没变。”
- “记忆细胞不是简单地’多’,它的’敏感’其实是因为它本来就处于半激活状态,所以一遇到抗原就能立刻反应。”
学生听到这些点,对免疫的理解就从”现象”进入了”机制”。



近期评论