


 扫码安装网站APP(Android版)
扫码安装网站APP(Android版)
来源公众号:一只蜉蝣的日常
siRNA的产生
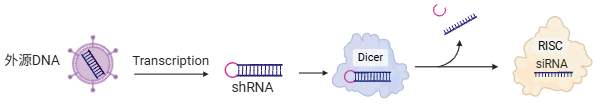
外源性来源
这是 siRNA 最常见的来源,也是实验室科研中最常用的手段。
- 化学合成:直接通过化学手段合成 21-23nt 的双链 siRNA,通过转染送入细胞。
- 体外转录:直接将长链dsRNA导入细胞
- 载体表达: 将能产生siRNA前体的DNA片段转入细胞,让细胞自己转录出shRNA。
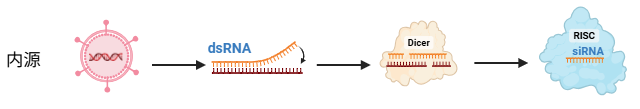
内源性来源
细胞内也会产生天然的 siRNA(称为 endo-siRNA-主要存在于卵子细胞和早期胚胎中)
- 病毒复制: 许多病毒(如轮状病毒)的遗传物质本身就是双链 RNA,在宿主细胞内复制时,会形成长链双链 RNA(dsRNA);有些单链 RNA 病毒,在宿主细胞内复制时,也会产生一个互补链,从而形成暂时的 dsRNA 状态。
- 基因组转录(如转座子):产生 endo-siRNA 或 shRNA-like 结构。
总结|siRNA产生机制
1、 触发
(1)dsRNA(双链RNA)
由两条完全独立的互补 RNA 单链通过碱基配对结合而成。
(2)shRNA(短发卡 RNA)
一根单链RNA,但序列设计成了“互补—回路—互补”的模式。这根单链会像发卡一样折叠,中间由一个不配对的环(Loop)连接,形成类似发夹的结构。
2、加工
一个叫Dicer的“剪切机”将长链RNA切成大约21个碱基对的双链RNA duplex的小片段,这就是siRNA。(未来与mRNA互补的链是引导链,另外一条是随从链)
3、组装
双链siRNA被装载到AGO2等蛋白上,形成前体RISC(RNA诱导的沉默复合体),随后,双链解开,随从链被丢弃,引导链被保留,形成成熟的、有活性的RISC。
备注:1、AGO2蛋白在哺乳动物中,四种AGO蛋白(AGO1-4)中,只有AGO2具有核酸内切酶活性,是切割mRNA的关键执行者。2、AGO2蛋白会选择双链中一条热力学稳定性较低的链作为成熟的引导链,另一条随从链则被降解并排出
4、执行
RISC复合体在细胞中巡逻,siRNA像“导航”一样,通过碱基配对,精准找到与之完全匹配的靶标mRNA(基因的“信使”)。然后,AGO2蛋白像剪刀一样将mRNA剪断、降解,从而阻止这个基因生产蛋白质,实现基因沉默。
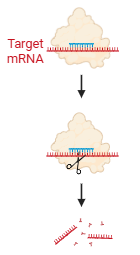

RNAi的调控机制
RNA 干扰(RNA interference, RNAi)是真核生物中由小干扰 RNA( siRNA)所介导的基因沉默现象。
RNAi 在细胞质中沉默基因表达的手段主要是通过 siRNA-RISC 介导的核酸内切酶作用降解 mRNA。
01 siRNA介导的基因表达转录水平调控机制
RNAi机制不仅能作用于细胞质中的成熟mRNA,还能进入细胞核,在更上游的环节(染色质水平和转录过程)调控基因表达,这被称为核内RNAi。
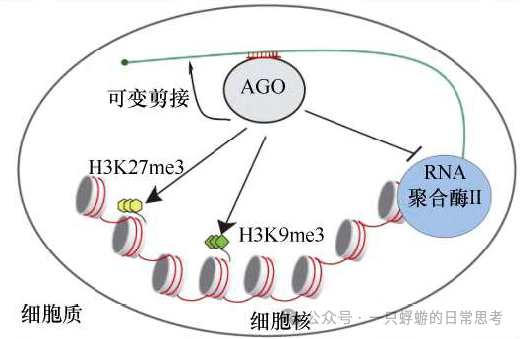
- 介导表观遗传修饰(染色质沉默):
- 机制:细胞核内的siRNA(或与其类似的途径)可以引导AGO蛋白及相关复合体定位到染色质上的同源DNA区域,进而招募组蛋白修饰酶或DNA甲基化酶,改变局部染色质状态。
- 组蛋白修饰:如诱导组蛋白H3第9位赖氨酸三甲基化或第27位赖氨酸三甲基化。这些是异染色质的标志,能抑制基因转录。DNA甲基化:在植物中尤为突出,存在RNA指导的DNA甲基化途径。
- 生物学功能:在酵母、植物、线虫等模式生物中,该机制对于着丝粒、端粒等异染色质区域的形成和维持、沉默转座子、以及发育基因的时序性调控至关重要。在哺乳动物中,核内RNAi的作用仍在深入研究。
- 影响转录过程本身:
- 抑制转录延伸:在线虫等生物中,核内siRNA/AGO复合体(如NRDE-3)可结合到正在转录的pre-mRNA上,进而招募蛋白导致RNA聚合酶II发生暂停或提前终止,从而阻碍转录的完成。
- 双向调控:某些AGO蛋白(如线虫的CSR-1)结合的小RNA反而能促进靶标基因的转录,其机制尚不完全清楚,提示了核内小RNA调控的复杂性。
- 参与其他核内过程:
- 调控RNA选择性剪接:在哺乳动物细胞中,siRNA或dsRNA可以将AGO蛋白招募到特定pre-mRNA上,改变其剪接方式,从而产生不同的蛋白质亚型。这一过程不依赖于mRNA的降解。
- 维持染色体稳定性:RNAi组分参与染色体配对和分离。例如,在裂殖酵母和线虫中,RNAi机器对于着丝粒功能和有丝分裂/减数分裂中染色体的正确分离是必需的。
02 RNAi引起的毒副作用
siRNA的脱靶效应
- siRNA通过与靶基因RNA完全互补配对来实现特异性沉默(称为“靶上效应”)。然而,如果siRNA与非靶基因的RNA发生部分互补配对(特别是其第2至第7位碱基与靶RNA互补),就会以类似于microRNA的机制,抑制该非靶基因mRNA的翻译并加速其降解。这种非特异性抑制靶基因以外其他基因表达的现象,就称为脱靶效应。
- 后果:
- 功能误读:在科学研究中,siRNA可能会影响几十甚至几百个基因的表达,导致对靶基因功能的错误解读,产生假阳性结果。
- 多重来源:
- 引导链脱靶:被RISC复合体保留的siRNA引导链可能与非靶mRNA部分配对。
- 随从链脱靶:在RISC装载过程中,本应被丢弃降解的随从链也可能被保留,并产生自身的脱靶效应。
- 蛋白依赖差异:在哺乳动物中,AGO2是唯一具有切割活性的AGO蛋白,能高效介导完全互补的靶向切割。但AGO1、AGO3、AGO4也能结合siRNA,它们主要介导部分互补配对引发的翻译抑制,其介导脱靶效应的效率甚至可能比AGO2更高。
- 特殊风险:许多基因存在天然的反义链转录本。当使用siRNA研究此类基因时,其引导链沉默正义链RNA,而其随从链可能沉默反义链RNA,导致实验假象。
siRNA对内源miRNA的竞争抑制
- 不论是化学合成的siRNA,还是由载体表达的shRNA,其最终发挥作用都需要占用细胞内的核心加工机器和效应蛋白,这与内源miRNA的加工和功能通路存在直接竞争。
- 竞争资源:
- 加工因子:载体表达的shRNA会与内源miRNA前体竞争Drosha/DGCR8复合体、核质转运蛋白Exportin-5以及Dicer酶。
- 效应蛋白:所有进入细胞的siRNA(无论是外源导入还是内源产生)都会与内源miRNA竞争有限的AGO蛋白,以装载形成RISC复合体。
- 后果:这种竞争会导致细胞内源miRNA的加工和功能普遍受到抑制,引起其靶基因表达水平异常上调,破坏细胞正常的调控网络,从而导致细胞毒性。文档明确指出,在小鼠肝脏中长期高表达shRNA可因竞争Exportin-5和AGO2而导致肝损伤甚至死亡。
总结:RNAi的毒副作用主要源于其作用机制与非靶基因的交叉反应(脱靶),以及其与细胞内天然调控系统(miRNA通路)对有限细胞资源的争夺(竞争抑制)
 end
end
来源网址:表观遗传学:siRNA和RNAi



近期评论