

 扫码安装网站APP(Android版)
扫码安装网站APP(Android版)
作者:黄建华 朱 燕
摘要 从强光防御、强光保护、光系统Ⅱ(PSⅡ)损伤修复3道防线介绍了避光运动、紫外辐射屏蔽、状态转换、调整捕光天线大小、非光化学猝灭、循环电子传递、水-水循环、叶绿体呼吸、光呼吸、消除活性氧(ROS)、PSⅡ的降解和组装等保护机制。
光为植物光合作用提供能量,充足的光照是植物健康生长的基础,但过量光照会使光合生物细胞产生以活性氧(ROS)为主的有害产物,损伤光系统,引起光合作用降低,这是典型的光抑制现象。植物光反应主要由位于叶绿体类囊体膜上的光系统Ⅱ(PSⅡ)、细胞色素b6f 蛋白复合体(Cytb6f)、光系统Ⅰ(PSⅠ)和ATP合酶等协同完成。过量光照时,PSⅡ中的叶绿素作为分子氧的光敏剂,通过能量转移形成1O2,PSⅠ反应中心将电子直接转移给O2。1O2是强光下产生的主要ROS形式,1O2损伤PSⅡ超级复合体中的D1亚基从而导致光抑制。PSⅡ是光抑制发生的主要部位,但在低温等逆境条件下也会导致PSⅠ发生光抑制。此外,强光中的紫外线使叶绿素a(Chla)的构象及与蛋白间的位置发生改变,导致PSⅡ损伤,研究表明叶绿体中约1/3的PSⅡ损伤是由紫外线辐射造成的。为了避免光抑制,植物进化出一套光保护机制。本文系统梳理了植物光保护机制。
1 第1道防线——强光防御
第1道防线主要是通过调整光的吸收来防御强光。在强光照下,植物可通过避光运动、紫外辐射屏蔽、类囊体膜的状态转换及调整捕光天线大小等避免吸收更多的光能。
1.1 避光运动 植物可通过叶片和叶绿体的运动来避免强光伤害。当光照过强时,植物叶片与太阳光线平行,减少吸收太阳辐射;同时叶绿体中的光受体PHOTO2感受光照强度,使叶绿体沿着平行于光线的方向分布,减少光能的吸收。
1.2 紫外辐射屏蔽 紫外线会促进叶表皮细胞酚类化合物(黄酮醇、花青素等)的合成和积累,酚类化合物对紫外线有屏蔽作用,减少紫外线对PSⅡ的损伤。
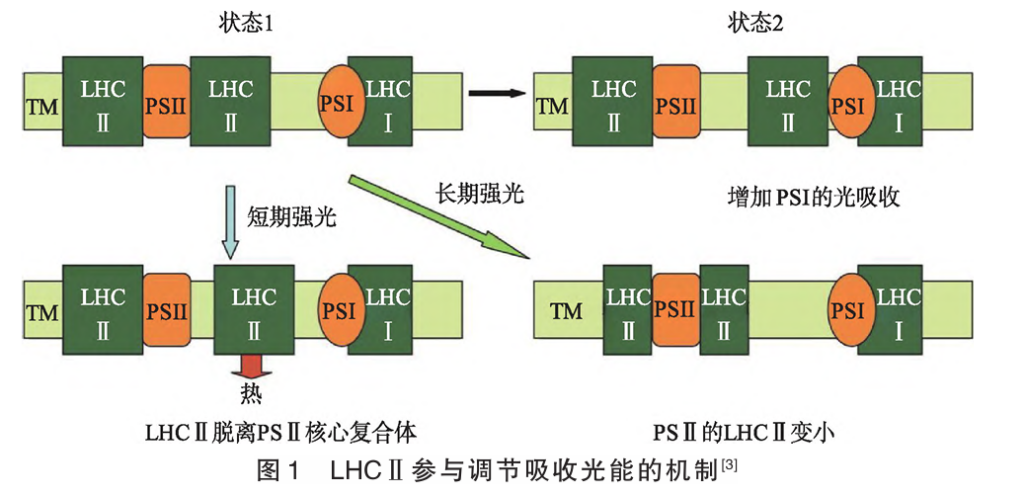
1.3 类囊体膜的状态转换及调整捕光天线大小 PSⅠ和PSⅡ分别由各自的核心复合物和LHC 组成。PSⅠ的核心结合捕光天线Ⅰ(LHCⅠ),形成PSⅠ-LHCⅠ复合物。PSⅡ的核心结合捕光天线复合物Ⅱ(LHCⅡ),形成PSⅡ-LHCⅡ复合物。植物中PSⅠ-LHCⅠ相对稳定,而PSⅡ-LHCⅡ的LHCⅡ与核心复合物的结合有紧密结合、中等强度结合和松散结合多种形式,可根据光照强度进行调节。当PSⅡ-LHCⅡ相对于PSⅠ-LHCⅠ被过度激发时,部分LHCⅡ被磷酸化,从PSⅡ-LHCⅡ上解离下来并结合到PSⅠLHCⅠ上,形成PSⅠ-HCⅠ-LHCⅡ型复合物,平衡激发能在2个光系统间重新分配,如图1所示,状态1向状态2的转换,预防PSⅡ-HCⅡ过度激化。当状态1向状态2转换不能实现光防御时,短期强光通过部分LHCⅡ与PSⅡ核心复合物分离减少光能吸收,长期强光通过缩小LHCⅡ减少光能吸收,见图1。
2 第2道防线——强光保护
第2道防线主要是通过非光化学猝灭(NPQ)、循环电子传递、水-水循环、叶绿体呼吸、光呼吸、消除ROS等对光系统进行保护。
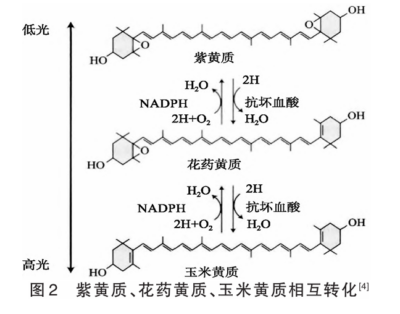
2.1 NPQ 植物光合色素吸收的光能有3个去向,即用于光合作用、以热能散失、以荧光的形式发光。通过光化学途径猝灭叶绿素荧光称为光化学猝灭(qP),通过光化学途径之外的途径猝灭叶绿素荧光称为非光化学猝灭。叶黄素循环依赖的猝灭(qE)在NPQ中起着重要作用。叶黄素包含紫黄质、花药黄质、玉米黄质等,在强光照下,类囊体腔的pH迅速降低,PSⅡ的PsbS(光系统Ⅱ亚基S)活性增强,激活紫黄质脱环氧化酶,将紫黄质依次转化为花药黄质、玉米黄质,玉米黄质结合到捕光天线蛋白上,引起其结构变化,导致热能耗散;当光强减弱时,过程发生逆转,见图2。
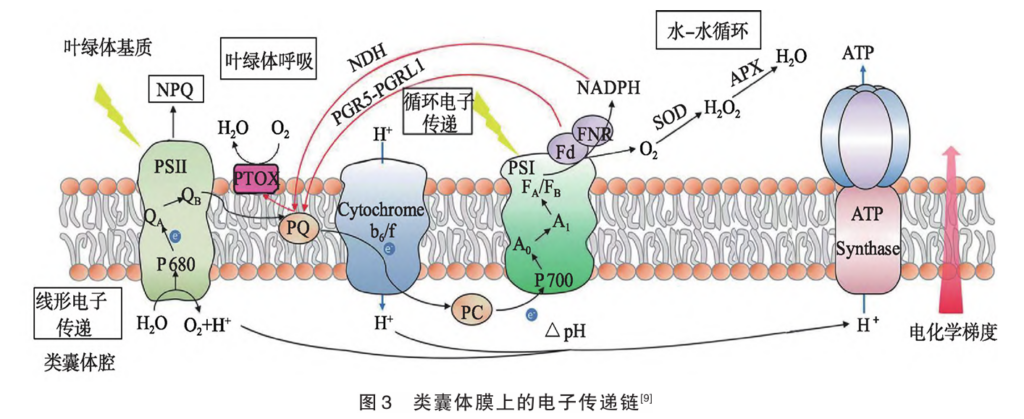
2.2 循环电子传递、水-水循环、叶绿体呼吸 光合作用放氧复合体裂解水产生的电子的传递途径分为线形电子传递途径(LEF,电子依次经过PSⅡ、Cytb6f、PSⅠ,最后传给 NADP+形成NADPH)和循环电子传递途径[CEF,传到PSⅠ的电子经质体醌(PQ)、Cytb6f又回到PSⅠ]。典型的循环电子传递有PGR5/ PGRL1介导途径和NDH介导途径(图3)。在强光下,PSⅠ介导的循环电子传递能向类囊体腔中运入H+,增强类囊体膜上△pH的生成。一方面诱导qE耗散过多光能提高电子传递效率[5];另一方面驱动合成更多的ATP,提高ATP/NADPH比值,促进卡尔文循环(卡尔文循环固定CO2需要ATP/NADPH的比值是1.5,而LEF生成ATP/NADPH的比值一般为1.3),从而调节LEF电子传递速度,提高放氧速率,保护PSⅡ。强光下,PSⅡ水光解产生的电子供给PSⅠ后,部分用于O2的还原,生成O2,O2在超氧化物歧化酶(SOD)催化下生成H2
O2,H2O2在抗坏血酸氧化物酶(APX)催化下生成H2O,这一过程称为水-水循环[6]。水-水循环一方面直接保护PSⅠ;另一方面增强类囊体膜上△pH的生成,诱导qE并提升ATP/NADPH比值,加快卡尔文循环过程,调节LEF电子传递速度,保护PSⅡ。经NDH介导途径传给PQ的电子在质体末端氧化酶(PTOX)的作用下传给分子氧生成水,这一过程称为叶绿体呼吸。叶绿体呼吸可防止PQ在强光条件下过度还原,减轻光损伤过程中对PSⅡ修复的抑制作用。
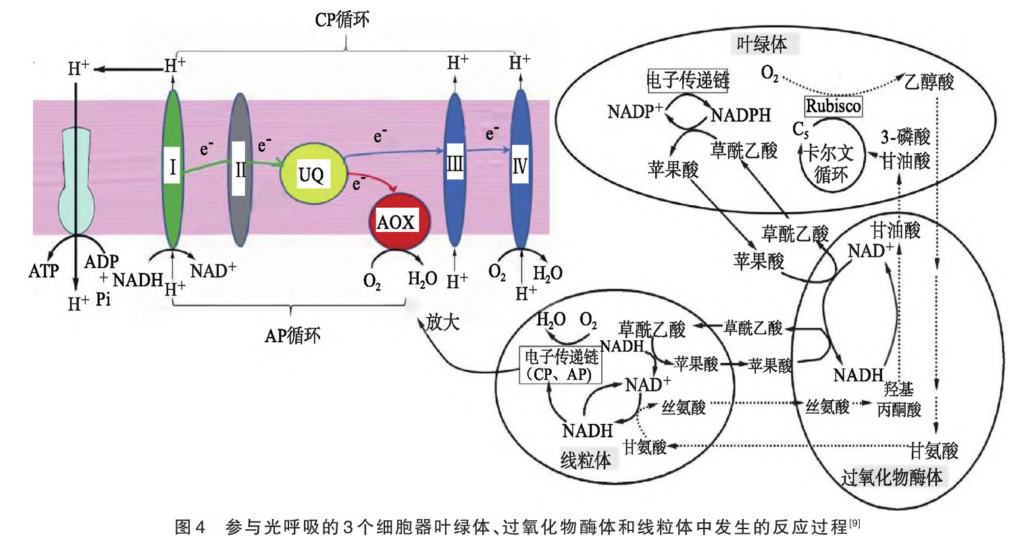
2.3 光呼吸 正常条件下,光合作用光反应产生的NADPH要多于暗反应消耗的NADPH,过剩NADPH可通过苹果酸-草酰乙酸穿梭转移至线粒体。Rubisco既可催化RuBP的羧化反应,也可催化RuBP的加氧反应,强光下O2与CO2分压比升高,Rubisco催化RuBP的加氧反应产生磷酸乙醇酸,再经过氧化物酶体、线粒体产生甘油酸和CO2,具体过程见图4。其中线粒体电子传递链有细胞色素途径(CP)和交替氧化酶途径(AP),CP与ATP合成偶联,AP不与ATP合成偶联。由图4可知,强光下光合作用产生的过剩NADPH通过草酰乙酸-苹果酸穿梭进入过氧化物酶体参与光呼吸途径,经一系列反应后以甘氨酸的形式进入线粒体并转化为丝氨酸,将还原氢转移至线粒体。其中,一部分通过AP消耗以热能形式释放;另一部分通过草酰乙酸-苹果酸穿梭再进入过氧化物酶体参与光呼吸途径,经一系列反应最终还是转移到线粒体中通过AP消耗以热能形式释放,减少自由基的产生,既保护光系统,又防止PSⅠ、PSⅡ之间的电子载体过分还原,确保电子在电子传递链上正常传递。同时光呼吸产生的甘油酸和CO2可加快卡尔文循环,加快NADPH的消耗,从而实现对光系统的保护。
2.4 消除ROS 为了有效避免体内ROS的过度积累,叶绿体可通过谷胱甘肽还原酶、超氧化物歧化酶、过氧化物酶、过氧化氢酶、抗坏血酸、维生素E、类胡萝卜素等有效清除ROS,减弱对光系统的抑制。
3 第3道防线——PSⅡ损伤修复
若第1、2道防线不能有效避免强光对光系统的损伤,植物会启动第3道防线——PSⅡ损伤修复。强光胁迫后,PSⅡ核心蛋白磷酸化,导致PSⅡ复合物从基粒类囊体迁移到基质类囊体并解组装、降解,引起D1蛋白损伤,受损的D1蛋白需要不断降解和新的D1从头合成才能维持PSⅡ的活性,Deg蛋白酶和FtsH蛋白酶参与受损D1蛋白的降解。受损的PSⅡ复合物降解后,新合成的D1蛋白等PSⅡ成员蛋白发生再加工,重新组装成
有活性的PSⅡ复合物。强光下加快PSⅡ复合物受损蛋白(如D1蛋白)的降解从而提高新蛋白的合成、组装,实现强光的保护。
4 小结
植物强光保护的强光防御(避免过多光能的吸收)、强光保护(避免ROS的损伤)、PSⅡ损伤修复3道防线共同作用,维持较为稳定的光合功能。除了上述保护机制外,光合植物还会通过调整激素比例、叶绿体自噬(选择性自噬过度损伤的叶绿体)、叶绿体转化等实现强光下对自身的保护,相信随着研究的深入,将有更多的光保护机制被发现。提高植物光合能力是提高植物生产力的有效途径,自然强光不可避免地会导致植物电子传递和固碳能力降低,避免或缓解强光胁迫对光合作用的影响具有十分重要的意义。因此,如何激活光保护机制,高效表达光保护相关基因,以及导入外源光保护相关基因等对提高植物光合速率、提高作物产量具有重要意义,为实现植物安全高效优质栽培和作物育种改良提供理论基础。
来源:黄建华,朱燕.植物光保护机制的解析[J].生物学通报,2025,60(07):88-91.
来源网址:植物光保护机制的解析



近期评论