


 扫码安装网站APP(Android版)
扫码安装网站APP(Android版)
陈绍洲
广东省海丰县彭湃中学
摘要:生物的性别决定是“基因在染色体上”的基石。科学家在性别遗传的实验研究和生殖细胞的显微研究中,发现特殊的限性遗传与“基因在染色体上”存在某种必然的联系,从而将孟德尔遗传学说与染色体学说完美结合在一起。其中,摩尔根在其经典实验基础上通过果蝇红、白眼和雌、雄性别2对相对性状的自由组合假设,以遗传实验的方式证明了细胞学中性染色体决定性别的遗传机制。这一机制的揭示是遗传学历史上具有里程碑意义的重大事件。
1 生物学教材“基因在染色体上”的简述
“基因在染色体上”是人教版高中生物学教材必修2《遗传与进化》第2章第2节的内容。其内容引入了摩尔根(Thomas Hunt Morgan,1866—1945)果蝇的限性遗传的实验和对生物性别决定简洁的描述“生物学家在一些昆虫的细胞中发现了性染色体”。果蝇的体细胞中有4对染色体,3对是常染色体,1对是性染色体。在雌果蝇中,这对性染色体是同型的,用XX表示;在雄果蝇中,这对性染色体是异型的,用XY表示。由于白眼的遗传和性别相关联,而且与X染色体的遗传相似,于是,摩尔根及其同事设想,控制白眼的基因W在X染色体上,而Y染色体上不含有它的等位基因。这样的描述很大程度上基于现代染色体学说已经建立以后对生殖细胞显微研究的认识,但性别遗传实验的科学史背景是比较复杂的。为尊重科学史实,结合1910年摩尔根等科学家对性别的认识,对相关问题进行更严谨的分析。
2 1910年前科学家对生物性别认识的简介
因光学显微镜和染色技术的发展,科学家对生殖细胞的显微研究有了一定的认知。1891年,亨金(Hermann Henking,1858—1942)在雄性无翅红蝽(Pyrrhocoris apterus)精子中观察到11对和1条单独的染色体,这条单独的染色体只进入一半数量的精子中,他将这个未知的染色体在图上标记为字母“X”[1]。但不知道这条染色体的作用,仅仅是一个符号,这也是“X染色体”概念的由来。1901年,麦克朗(Clarence Erwin Mc Clung,1870—1946)提出副染色体(accessory chromosome)决定性别的假说,首次提出副染色体与生物性别的关系[2]。1905年,威尔逊(Edmund Beecher Wilson,1856—1939)提出昆虫的性别决定存在2种类型(分别称为A和B)的假说。A类昆虫的雌性有1对大小相等的副染色体,雄性只有1条副染色体;B类昆虫的雌性同样有1对大小相等的副染色体,雄性有1条与雌性相等大小的副染色体和1条较小的副染色体[3]。1906年,威尔逊首次使用“性染色体(sex chromosome)”一词代替副染色体[4]。1909年,威尔逊首次在雄性昆虫中使用“X”和“Y”来区分副染色体[5],即性染色体。A、B类型昆虫雌性的性染色体组成用“XX”表示,A类型雄性的性染色体组成用“X”表示,而B类型雄性的性染色体组成用“XY”表示。威尔逊提出的性别决定学说基于当时对生殖细胞广泛的显微研究,有近百种昆虫的性染色体被观察,其中一群美国细胞学家处于领先地位[5],但没有在性别遗传的实验研究中得到证明。史蒂文斯(Nettie Maria Stevens,1861—1912)曾表示“决定性别的因素是否为孟德尔(Gregor Johann Mendel,1822—1884)性状的唯一希望似乎在于育种实验,而细胞学研究可能显示其形式是有利的”[6]。
性别遗传实验证明,细胞中性染色体与孟德尔遗传性状相关的实验就是摩尔根的果蝇限性遗传。摩尔根认为雄果蝇不存在Y染色体,野生雌性红眼用RRXX表示,突变雄性白眼用WWX表示,其中R与W分别表示红、白眼因子,X是性因子[7]。这种表示方法与教材相差甚远,却是当时最符合科学家认知水平的科学假设,主要原因是摩尔根受1910年前醋栗尺蠖(Abraxas grossulari‐ata)性别遗传实验的假设影响。
3 醋粟尺蠖杂交实验
1903年,雷纳(Raynor)和唐卡斯特(Leonard Doncaster,1877—1920)获得醋栗尺蠖(图1)及其乳白色变种(flavofasita)并进行杂交实验。乳白色变种是一种罕见的雌性品种,黑色斑点减少,呈部分线性,类似于白化病,其中黑色色素持续减少,但并非完全不存在。

图1 醋栗尺蠖
乳白色雌性与正常黑色雄性杂交,所有的后代都是正常黑色。然而,当杂种子一代的雌、雄个体进行交配,子二代中乳白色全是雌性,雄性没有出现乳白色,其他雌、雄个体都是正常黑色。雷纳和唐卡斯特通过多年大量的杂交实验证明变种乳白色是一种全新类型的孟德尔隐性遗传,由于只在雌性身上发生,因此进行了更精确的实验研究[8]。醋栗尺蠖的实验时间在1905—1910年,遗传学家和细胞学家对其生物性别的推测很可能占据首要位置[9]。这一奇特的孟德尔遗传现象让科学家隐约意识到醋栗尺蠖这一对相对性状可能与性别存在某种必然的联系。科学家提出至少4种假设解释性别和该性状的关系,试图突破细胞学和遗传学的距离。
雷纳和唐卡斯特认为,每种受精卵在性别上都是杂合的,并且每种性别都会产生雄配子和雌配子。假设:1)在F1中雌性个体形成配子存在耦合,即雄性卵细胞都带有黑色因子,雌性卵细胞都带有乳白色因子。2)F1中雄性个体形成配子没有耦合。3)受精结合只能发生在具有相反性别的配子之间。4)受精卵发育由卵细胞的雌、雄决定[8]。其遗传分析如图2所示。
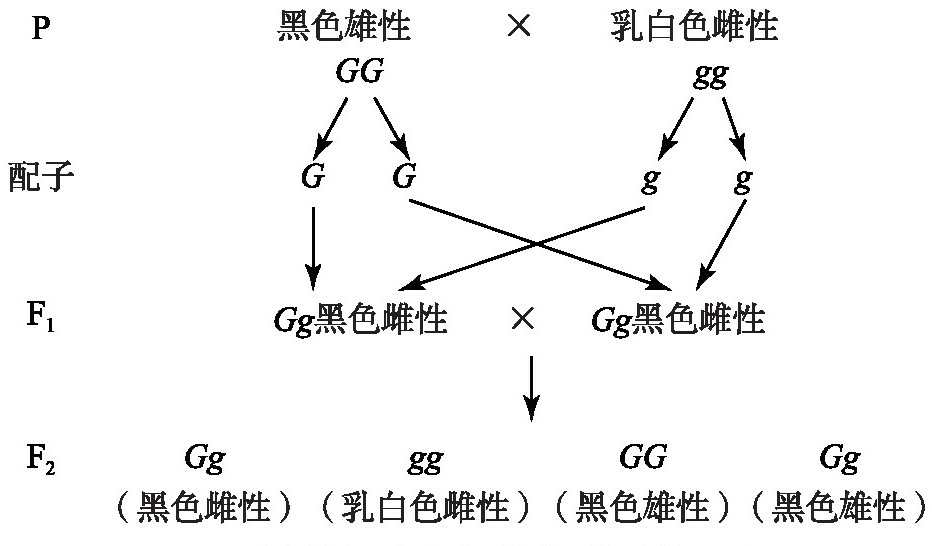
图2 雷纳和唐卡斯特假说遗传图解
庞尼特(Reginald Crundall Punnett,1875—1967)与贝特森(William Bateson,1861—1926)优化了唐卡斯特的假说:1)一只雌性在性别上是杂合的,雌性是显性的,雄性是纯合隐性的。2)在F1中,当2个显性因素,雌性的♀(♀表示雌性因子,且为显性的作用;♂表示雄性因子)和黑色(用G表示)因子共存时,它们之间存在互斥作用[10]。其遗传分析如图3所示。
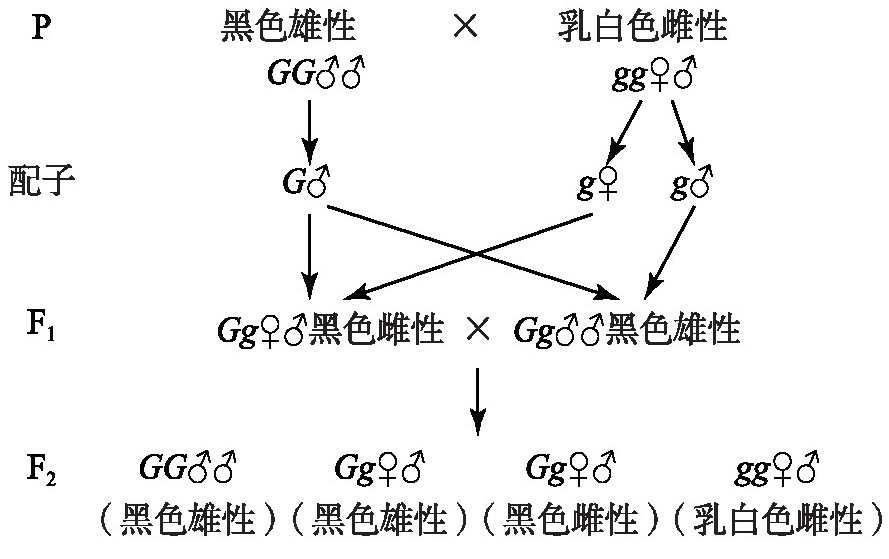
图3 庞尼特与贝特森假说遗传图解
卡斯特(William Ernest Castle,1867—1962)的假说:一条X符号代表雌性,没有X符号代表雄性,前提是采用贝特森的互斥假设(F1Gg X黑色雌性的G与性因子X互斥)[9]。结果可以得到解释,其遗传分析如图4所示。但是没有直接找到醋栗尺蠖雌性只有一条X性染色体、雄性没有X性染色体的证据。
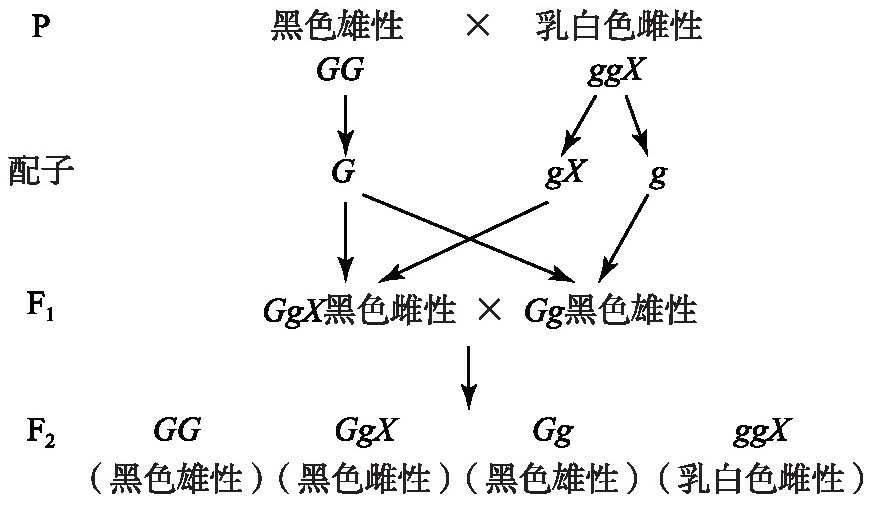
图4 卡斯特假说遗传图解
威尔逊的假说:雌性在性别上存在XY染色体,雄性存在YY。黑色G与乳白色g的染色体配对的同时,G与Y染色体总是联合连接[11]。其遗传分析如图5所示。
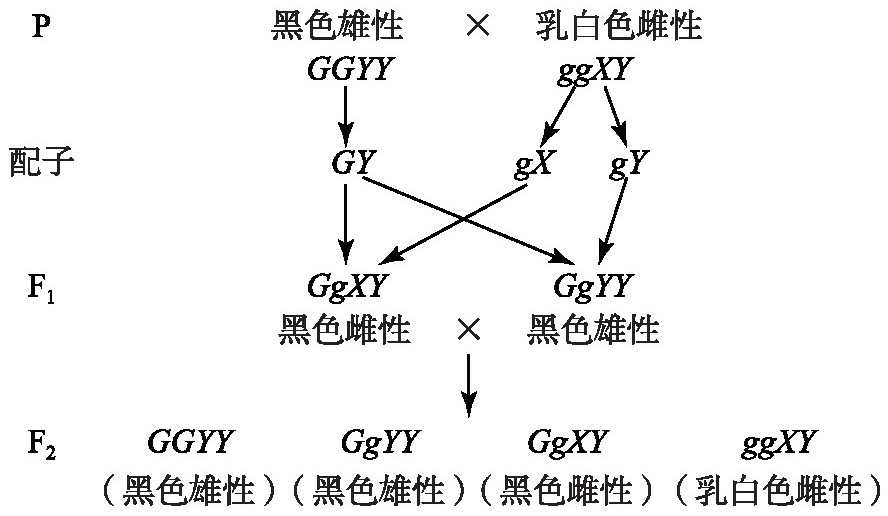
图5 威尔逊假说遗传图解
威尔逊作为细胞学家知道醋栗尺蠖细胞学研究的重要性。醋栗尺蠖精母细胞分裂呈现正常的外观。精母细胞的分裂,无论是杂交形式还是自交形式,都显示出非常复杂和令人困惑的现象,与其他昆虫中描述的任何现象都不同,无法在细胞学研究中找到雄性醋栗尺蠖存在2条相同Y染色体的证据。
4果蝇白眼突变的杂交实验
1910年,摩尔根进行了果蝇白眼突变的杂交实验[7],发现果蝇白眼突变与醋栗尺蠖的实验现象相反,只获得了雄性白眼突变。其遗传分析如图6所示。
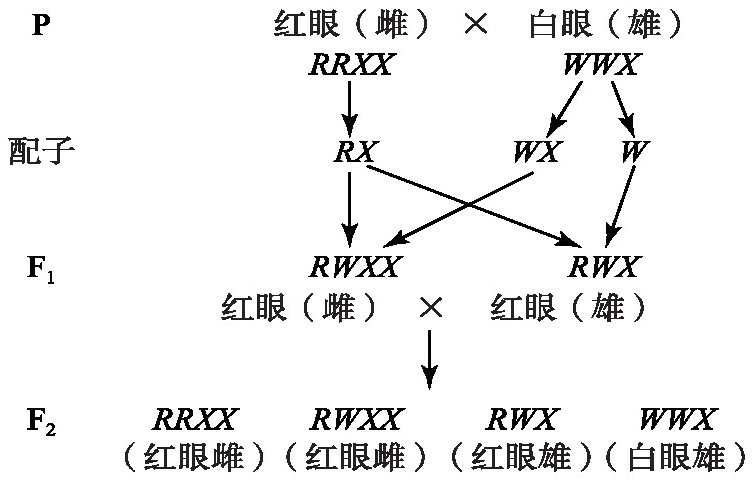
图6 果蝇杂交实验遗传图解
果蝇个体小,易饲养;在室温下10多天繁殖一代,一只雌果蝇一生能产生几百个后代,因此,摩尔根可以进行大量的实验进行验证。
摩尔根通过严谨实验分析果蝇性别和性状的遗传发现,野生雄性的2个性状(眼色和性别)都是杂合子RWX,而野生雌性(RRXX)是纯合子。野生雌性醋栗尺蠖在颜色和性别上都是杂合子,这是果蝇限性遗传的另一个佐证。
5 科学史实与教材的表述
摩尔根早期采用遗传分析果蝇限性遗传是一种探索,采用孟德尔自由组合定律分析2对相对性状(眼色和性别)。摩尔根根据当时果蝇细胞染色体组成的研究成果,认为雄果蝇只有一条X性染色体[12]。此时,摩尔根还没有将基因定位于染色体上的想法,所以采用X因子代替X染色体。这个假设的后果引起一个不能自圆其说之处——假设野生红眼雄性果蝇杂合子是RWX,不是RRX。摩尔根又发现了果蝇的短翅和黄身2种限性遗传的现象[12],进一步肯定,这些控制限性遗传的基因位于X染色体上。摩尔根意识到性别由性染色体决定。1914年,布里奇斯(Calvin Blackman Bridges,1889—1938)观察到雌性果蝇携带2条X染色体,雄性携带不相等的X-Y对,且Y染色体比X染色体大12%,雄性的Y精子代替“无X”的精子[13],这是摩尔根团队首次证明雄性果蝇存在XY染色体。
在摩尔根团队集中精力研究果蝇遗传实验和细胞学染色体组成期间,其他科学家在其他昆虫中发现新的性染色体组成。塞勒(Karl Gottlieb Seiler,1873—1961)发现红宝石虎蛾(Phragmatobia fuliginosa)雌性存在一对异形染色体[14],唐卡斯特发现醋栗尺蠖雌性也存在一对异形染色体[15]。
1915年,摩尔根团队尝试用染色体理论来解释孟德尔遗传现象,是遗传学历史上的一个里程碑。其中对果蝇性染色体的组成进行总结性的描述[16]。这就是今天教材所采用的“红眼雌性XWXW,白眼雄性XwY”。醋栗尺蠖雌性因存在一对异形染色体,摩尔根团队首次使用W和Z符号区分果蝇雌性同形性染色体,随后确定了不同生物存在不同的性别决定机制。
细胞学中性染色体决定性别的遗传机制的揭示是遗传学历史上具有里程碑意义的重大事件。
参考文献
- [1]Henking H. Untersuchungenüber die ersten entwicklungsvorgänge in den eiern der insektenⅡUber spermatogenese und beziehung zur eientwickling bei pyrrhocoris apterus[J].Zeitschrift für wissenschaftliche Zoologie,1891,51:685.
- [2]McClung C E. Notes on the accessory chromosome[J].Anatomical Anzeiger,1901,20:220.
- [3]Wilson E B. The chromosomes in relation to the determination of sex in insects[J].Science,1905,22:500.
- [4]Wilson E B. Studies on chromosomes.Ⅲ. The sexual differences of the chromosomegroups in Hemiptera,with some considerations on the determination and inheritance of sex[J].Journal of Experimental Zoology,1906,3(1):1.
- [5]Wilson E B. Recent researches on the determination and heredity of sex[J]. Science,1909,29:53.
- [6]Stevens N M. A study of the germ cells of certain Diptera,with reference to the heterochromosomes and the phenomena of synapsis[J]. Journal of Experimental Zoology,1908,5:359.
- [7]Morgan T H. Sex-limited inheritance in Drosophila[J].Science,1910,812(7):122.
- [8]Doncaster L,Raynor R G H. On breeding experiments with Lepidoptera[J]. Journal of Zoology,1906,76:125.
- [9]Morgan T H. Chromosomes and heredity[J]. The American Naturalist,1910,8:449.
- [10]Punnett R C,Bateson W.The heredity of sex[J].Journal of Genetics,1908,3:785.
- [11]Wilson E B. Secondary chromosome-couplings and the sexual relations in Abraxas[J].Science,1909,4:704.
- [12]Morgan T H. An attempt to analyze the constitution of the chromosomes on the basic of sex-limited inheritance in Drosophila[J].Journal of Experimental Zoology,1911,11:365.
- [13]Bridges C B. Direct proof through non-disjunction that the sex-linked genes of Drosophila are borne by the X chromosome[J]. Science,1914,50:107.
- [14]Seiler J. Geschlechts chromosome bei Lepidopteren[J].Zoologischer Anzeiger,1913,41:246.
- [15]Doncaster L. On the relations between chromosomes,sex-limited transmission and sex-determination in Abraxas grossulariata[J].Journal of Genetics,1914,6(4):1.
- [16]Morgan T H,Sturtevant A H,Muller H J,et al. The mechanism of Mendelian heredity[M]. New York:Henry Holt and company,1915:48.
来源:陈绍洲.性别决定与“基因在染色体上”的探究历程[J].生物学通报,2026,61(2):91-94.



近期评论